Faslılarla ilgili genetik araştırmalar - Genetic studies on Moroccans
Fas genetiği halkının genetik tarihini kapsar Fas ve bu soyun dünya popülasyonları üzerindeki genetik etkisi. Coğrafyadan büyük ölçüde etkilenmiştir.
Tarih öncesi çağlarda, güneyde Sahra çölü ve kuzeyde Akdeniz önemli coğrafi engellerdi. Batı Asya ve Kuzeydoğu Afrika, Süveyş'te tek bir kara kütlesi oluşturur. Mağrip ve Güneybatı Avrupa, Cebelitarık Boğazı'nda yalnızca 15 km (9 mil) ile ayrılır ve benzer bir mesafe, Afrika Boynuzu'nu Bab el-Mandeb boğazındaki Arap yarımadasından ayırır. Azami buzul dönemlerinde olduğu gibi deniz seviyesinin düşük olduğu dönemlerde, şu anda Akdeniz'de ve muhtemelen Cebelitarık boğazları arasında batmış olan adalar yaşanabilir olacaktı. Bunlar, Afrika ve Avrupa arasında çift yönlü demik yayılmayı teşvik etmiş olabilir. Sahra'nın daha yağışlı dönemleri sırasında, Orta Afrika'daki bazı popülasyonlar kuzeye, Kuzey Afrika'nın güney kısımlarına taşınmış olacaktı. Batı Asya nüfusu, Süveyş, Bab el-Mandeb veya Akdeniz yoluyla gelen ıslak Sahra'ya da çekilebilirdi.
Bu coğrafi etkilerin bir sonucu olarak, Fas nüfusunun genetik profili, değişken derecelerde Kuzeydoğu Afrika, Avrupa, Batı Asya ve Batı Afrika unsurlarının yanı sıra, otokton Maghrebi soylarının karmaşık bir mozaikidir. Kuzeybatı Afrika, çevredeki bölgelerden gen akışı yaşamış olsa da, bazı kısımlarda uzun süreli genetik izolasyona maruz kalmıştır. Bu, bazı Maghrebi popülasyonlarında, özellikle de Berberi konuşan bazı izole gruplarda, ayırt edici genetik belirteçlerin gelişmesine izin verdi.
Tarih öncesi ve antik dönem
Bugünkü Fas'ın o zamandan beri iskan edildiği düşünülüyordu. Paleolitik Bazen, MÖ 90.000 ile 190.000 arasında, ancak 300.000 yıllık bir homo sapien'in keşfinden sonra artık durum söz konusu değildir ve bunun yerine, ilk zamanlardan beri aynı kanıtlarla insanlar tarafından iskan edildiği öne sürülmektedir.[1] Sırasında Üst Paleolitik, Mağrip bugün olduğundan daha verimliydi, bir savana bugünün kurak manzarasından daha fazlası.[2] 22.000 yıl önce Aterian tarafından başarıldı Iberomaurusian İber kültürleriyle benzerlikler paylaşan kültür. Iberomaurusian arasında iskelet benzerlikleri gözlenmiştir. Mechta-Afalou cenaze törenleri ve Avrupa Cro-Magnon kalır. Fas'taki Iberomaurusian endüstrisinin yerini, Capsian kültürü.




Kuzey Afrika ve Fas yavaş yavaş ortaya çıkan Akdeniz dünya tarafından Fenikeliler Erken Klasik dönemde ticaret kolonileri ve yerleşim yerleri kuran.[3] Mogador, MÖ 6. yüzyılın başlarında bir Fenike kolonisiydi.[4][sayfa gerekli ]
Fas daha sonra, merkezi Kuzey Afrika imparatorluğunun bir parçası oldu. Kartaca. Bilinen en eski bağımsız Fas devleti, Mauretania Berberi krallığı kralın altında Bocchus I. Kuzey Fas'taki bu krallık, bugünkü durumuyla karıştırılmamalıdır. Moritanya, en az MÖ 110'a tarihlenir.[5]
Roma imparatorluğu Bu bölgeyi MÖ 1. yüzyıldan itibaren kontrol ederek, Mauretania Tingitana. Hıristiyanlık MS 2. yüzyılda tanıtıldı ve Roma kasabalarında din değiştirenler kazandı. köleler ve bazı Berberi çiftçileri.
MS 5. yüzyılda Roma İmparatorluğu'nun gerilemesi ile bölge ilk olarak kuzeyden Vandallar ve sonra Vizigotlar. MS 6. yüzyılda kuzey Fas, sözde Doğu Roma'nın bir parçasıydı veya Bizans imparatorluğu. Bu süre boyunca, Fas'ın iç kesimlerindeki yüksek dağlarda yaşayan Berberi sakinleri boyun eğmeden kaldı.
Erken İslam dönemi
670 yılında, ilk İslami fethi Kuzey Afrikalı kıyı düzlüğü altında yer aldı Uqba ibn Nafi altında görev yapan genel Emeviler nın-nin Şam. Emevi Müslümanlar dillerini, yönetim sistemlerini ve İslam'ı Fas'a getirdiler. Berberilerin çoğu, çoğunlukla Arap yönetimi geriledikten sonra yavaş yavaş İslam'a döndü. Modern Fas bölgesindeki ilk bağımsız Müslüman devlet, Nekor Krallığı, bir emirlik Rif Dağları. Tarafından kuruldu Salih ibn Mansur 710'da bir müşteri devleti olarak Rashidun Halifeliği. Salgınından sonra Büyük Berberi İsyanı 739'da Berberiler başka bağımsız devletler kurdular. Miknasa nın-nin Sijilmasa ve Barghawata.
Ortaçağ efsanesine göre, İdris ibn Abdallah Abbasiler'in Irak'taki kabilesini katletmesinden sonra Fas'a kaçmıştı. İkna etti Awraba Berberi kabileleri uzak Abbasi halifelerine bağlılıklarını kırmak için Bağdat ve o kurdu İdris Hanedanı 788 yılında. İdrisidler kuruldu Fes başkentleri ve Fas, Müslüman eğitiminin merkezi ve büyük bölgesel güç. Idrissidler 927'de Fatımi Halifeliği ve onların Miknasa müttefikleri. Miknasa, 932'de Fatımiler ile ilişkilerini kestikten sonra, Fatımiler tarafından iktidardan uzaklaştırıldılar. Maghrawa of Sijilmasa'nın 980.
Berberi hanedanları


11. yüzyıldan itibaren, bir dizi güçlü Berberi[6][7][8] hanedanlar ortaya çıktı. Altında Almoravid hanedanı [9] ve Almohad hanedanı Fas, Mağrip'e, günümüz İspanya ve Portekiz'in çoğuna ve Batı Akdeniz bölgesine hakim oldu. 13. ve 14. yüzyıllarda Merinidler Fas'ta iktidarı elinde tuttu ve ülkenin başarılarını çoğaltmaya çalıştı. Almohads Cezayir ve İspanya'daki askeri kampanyalarla. Onları takip etti Wattasidler. 15. yüzyılda Reconquista Orta ve güney İspanya'da Müslüman yönetimine son verdi ve Müslümanlar ve Yahudiler Fas'a kaçtı.[10] Portekizce 15. yüzyılda Atlantik kıyılarını kontrol etme çabaları, Fas'ın iç kesimlerini büyük ölçüde etkilemedi. Elizabeth Allo Isichei'ye göre, "1520'de bir kıtlık Fas'ta o kadar korkunçtu ki, uzun bir süre başka olaylarla tarihlendirildi. On altıncı ve on dokuzuncu yüzyıllar arasında Fas nüfusunun 5'ten 3 milyonun altına düştüğü öne sürüldü. "[11]
Şerif hanedanları
1549'da bölge, Arap hanedanlarının eline geçti. İslam peygamberi, Muhammed: ilk önce Saadi hanedanı 1549'dan 1659'a kadar hüküm süren ve sonra Alevi hanedanı, 17. yüzyıldan beri iktidarda kalan.

Saadi Hanedanlığı döneminde ülke geri püskürtüldü Osmanlı akınlar ve bir Portekizce işgali Ksar el Kebir savaşı 1578 yılında. Ahmed el-Mansur Saltanat'a yeni zenginlik ve prestij getirdi ve Batı Afrika'ya yapılan büyük bir sefer, Songhay İmparatorluğu 1591 yılında. Ancak, Sahra çok zor oldu. Mansur'un ölümünden sonra ülke oğulları arasında bölündü.
1666'da Fas, Alevi Hanedanı, o zamandan beri Fas'ın iktidarı olan. Fas, İspanya'nın saldırısıyla karşı karşıyaydı ve Osmanlı İmparatorluğu batıya doğru uzanıyor. Aleuiler pozisyonlarını sağlamlaştırmayı başardılar ve krallık bölgedeki öncekilerden daha küçükken oldukça zengin kaldı. Yerel kabilelerin muhalefetine karşı İsmail İbn Şerif (1672–1727) birleşik bir devlet yaratmaya başladı.[12] Jaysh d'Ahl al-Rif ile ( Riffian Ordu) ele geçirdi Tanca -den İngilizce 1684'te ve İspanyol itibaren Larache 1689'da.
Fas, yeni doğan Amerika Birleşik Devletleri'ni 1777'de bağımsız bir ulus olarak tanıyan ilk ülkeydi.[13][14][15][16] Başlangıcında Amerikan Devrimi Amerikan ticaret gemileri Atlantik Okyanusu tarafından saldırıya uğradı Berberi korsanları. 20 Aralık 1777'de Fas Sultanı Muhammed III Amerikan ticaret gemilerinin saltanatın koruması altında olacağını ve böylece güvenli geçiş sağlayabileceğini ilan etti. Fas-Amerikan Dostluk Antlaşması, 1786'da imzalanmış, ABD'nin en eski bozulmamış dostluğu olarak duruyor antlaşma.[17][18]
Genetik kanıt
Arasında gözlemlenen genetik yakınlık Faslılar ve Güney Avrupalılar her iki grubun da ortak bir atayı paylaşmasından kaynaklanmaktadır. Üst Paleolitik, Ve içinde Neolitik veya alternatif olarak, Güney İberya'da paylaşılan tarih sırasında Mağribi askerler.[19] Ocak 2012'de yayınlanan bir genetik çalışma, yerli halkın Kuzey Batı Afrika ata, en yakın popülasyonlarla ilişkili görünür. Afrika ama "farklılık Fas halkı ve Yakın Doğu /Avrupalılar muhtemelen öncesinde Holosen (> 12.000 ya) ve Paleolitik (> 40.000BC). "[20]
Son araştırmalar önemli olmadığını açıkça ortaya koyuyor genetik Arapça konuşan Faslı nüfus ile Arapça konuşmayan Faslı nüfus arasında farklılıklar vardır. İnsan lökosit antijeni HLA DNA veriler, çoğu Faslılar Arap olmayanların ikisi de etnolinguistik kimlik ve Arap etnolinguistik kimliğine sahip olanlar, Berberi köken ve şecere doğru Araplar Arabistan'dan sadece Fas'ı değil, geri kalanını da işgal eden Kuzey Afrika artı ispanya 7. yüzyılda gen havuzuna önemli ölçüde katkıda bulunmadı.[21][22] Mağribi mülteciler ispanya sahil kasabalarına yerleşti.[23]
Aslında, 2000 tarihli bir makaleye göre Avrupa İnsan Genetiği Dergisi, Kuzey-Batı Afrika'dan Faslılar genetik olarak İberler ikisinden de Sahra Altı Afrikalılar nın-nin Bantu etnik köken ve Orta Doğulular.[24] Sonuçlar ya Faslıların İspanyollarla tarih öncesi bir akrabalık ilişkisi olduğunu ya da Fas'tan İspanya'ya Mağribi (yani Berberi) girdisinin bir sonucu olduğunu ya da Fas'a sürgün edilen Müslümanlaştırılmış yerli İspanyollardan Fas'a genetik girdinin bir sonucu olduğunu gösteriyor. Katolik Reconquista'yı takiben İspanya'dan gelen Müslümanlar (burada Müslüman sürgünler, her halükarda nihai olarak esas olarak Fas Berberi kökenli olan yabancı kökenli Müslüman nüfus olan Mağribi nüfusun yanı sıra, çoğunluğu olmasa da, aslında yerli İspanyollar İslam'a dönüşüyor), bu da İspanyol DNA'sının Fas'a genetik girdisi ile sonuçlandı, Arapların Fas'ı fethi sırasında Arap soyundan gelen Arapların Fas'ta sahip olduğu herhangi bir genetik girdiden çok daha fazla.
Farklı olan lokus incelendiğinde arasında yakın benzerlik ortaya çıktı Berberiler ve esas olarak Faslı Arapça konuşanlardan oluşan diğer Kuzey Afrika grupları, mevcut Fas nüfusunun güçlü bir Berberi geçmişine sahip olduğu hipoteziyle uyumludur.[25]
Çeşitli popülasyon genetiği Gabriel Camps ve Charles-André Julien gibi tarihçilerle birlikte yapılan çalışmalar, modernin gen havuzunun büyük bir kısmının Kuzeybatı Afrikalılar dilsel gruba bakılmaksızın, Berberi İslam öncesi dönemin nüfusları.[26]
Göre X-Kromozom SNP analizleriyazarlar, Kuzeybatı Afrika'da berberler ve Araplar arasında yüksek bir genetik homojenlik olduğunu bildirdiler, bu nedenle, Araplaştırma bu bölgenin kültürel bir olguydu. yapmadı soy popülasyonunun değiştirilmesini ima eder. Elde ettiğimiz sonuçlar, kuzeybatı Afrika'nın erken yerleşiminin hipotezini desteklemektedir. Orijinal berber popülasyonu, çevredeki alanlardan düşük bir genetik akış almış gibi görünüyor. Fas nüfusunun genetik farklılaşmasını açıklamak için farklı hipotezler öne sürüldü. Başlangıçtaki bir genetik sapma, alel frekansı dağılımında, belirli bir coğrafi izolasyon seviyesi nedeniyle yeniden kurulmamış farklılıklara neden olmuş olabilir. Cebelitarık Boğazı, birçok yazar tarafından önemli bir genetik engel olarak tanımlanmıştır. Muhtemelen Kuzeybatı Afrika ile İber Yarımadası'nın güneyi arasında belirli bir düzeyde genetik değişim gerçekleşmiş olsa da, bu alanda keskin frekans değişiklikleri tanımlanmıştır. Ayrıca Sahra Çölü'nün Kuzeybatı Afrika popülasyonlarının Sahra Altı popülasyonlarından genetik izolasyonundan sorumlu olduğu ileri sürülmüştür. Neolitik demik yayılmanın Akdeniz bölgesindeki etkisi konusunda bir fikir birliği yoktur. Sonuçlarımıza göre, Neolitik genişlemelerin ve / veya sonraki göç olaylarının Kuzeybatı Afrika nüfusu üzerinde düşük bir etkisi olacaktı.X-Kromozom SNP analizleri
Fas Y-DNA kromozomu
E-M215
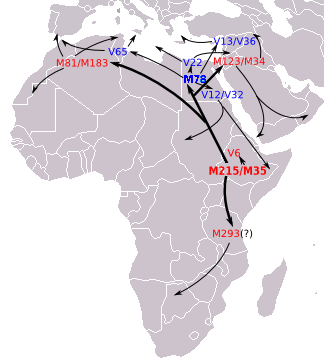
E1b1b (E-M215), Kuzey Afrika'daki en yaygın haplogruptur. E-M215 ve onun baskın alt sınıfı E-M35'in yaklaşık 22.400 yıl önce Doğu Afrika'da ortaya çıktığı ve daha sonra Kuzey Afrika'ya ve oradan da Batı Asya'ya dağılacağı düşünülüyor.[29][30] E1b1b1 clade şu anda çeşitli şekillerde bulunur Fas. Toplam E1b1b1 (E-M35) frekansları% 93,8'e ulaştı Faslılar.[31]
E-M215, bilinen tüm modern E-M215 erkeklerini, E-M35 ve E-M281'i içeren iki eski şubeye sahiptir. Bu ikisinin dışında yerli bir popülasyonda onaylanan tek dal, Etiyopya sırayla bilinen dört şubesi olan E-M35, E-V68, E-Z827, E-V6 ve E-V92. E-V68 ve E-V257 en yüksek sayılarda bulundu Kuzey Afrika ve Afrikanın Boynuzu; aynı zamanda daha düşük sayılarda Orta Doğu ve Avrupa ve izole popülasyonlarda Güney Afrika.
E1b1b aşağıdakiler arasında baskındır: Afro-Asya konuşmacıları. Dilsel grup ve E-M35 soyunun taşıyıcıları, yüksek olasılıkla birlikte ortaya çıkma ve dağılma olasılığına sahiptir. bu dil ailesinin menşe bölgesi Afro-Asya konuşma geçmişine sahip halklar arasında.[32][33]
E-M35'in tüm ana alt dallarının, ana sınıfla aynı genel alandan kaynaklandığı düşünülmektedir: Kuzey Afrika, Afrikanın Boynuzu veya yakınlardaki alanlar Yakın Doğu. E-M35'in bazı şubeleri binlerce yıl önce Afrika'yı terk etti. Örneğin, Battaglia vd. (2007) E-M78'in (bu makalede E1b1b1a1 olarak adlandırılır) Avrupa'da 10.000 yıldan uzun süredir bulunduğu tahmin edilmektedir.
E-M81

E1b1b1b1 (E-M81), vakti zamanında E1b1b1b, E3b1b, ve E3b2, en yaygın Y kromozom haplogrubudur. Fas, alt sınıfının hakim olduğu E-M183.
14.200 yıl önce Kuzey Afrika'da ortaya çıktığı düşünülüyor.[34] Ebeveyn sınıfı E1b1b (E-M215) İlk olarak Afrika Boynuzu'nda yaklaşık 42.600 yıl önce ortaya çıktığına inanılıyor.[35]
Bu haplogrup, ortalama% 85 frekansa ulaşır. Kuzey Afrika. Sıklığı, Saharawis dahil bazı Faslı Berberi popülasyonlarında yaklaşık% 80 veya daha fazla iken, Mısır'daki bu aralığın yaklaşık% 28'ine düşer.[30][36][37]
Bu Berberi grupları ve diğerleri gibi sınıfın yaygınlığı nedeniyle Mozabitler, Riffians, Chleuhs, Orta Atlas ve Kabyle bazen genetik olarak adlandırılır Berber işaretçisi.
The Berber haplogroup subclades'in bu filogenetik ağacı, YCC 2008 ağacına ve ISOGG tarafından özetlendiği gibi daha sonra yayınlanan araştırmaya dayanmaktadır.[38][39][40]
- E1b1b1b (L19, V257)
- E1b1b1b1 (M81)
- E1b1b1b1a (M107) Underhill vd. (2000).
- E1b1b1b1b (M183) Bu sınıf, E-M81'de son derece baskındır. Aslında Karafet vd. (2008) bunu E-M81'in bir alt sınıfı olarak tanımlamaya devam ediyor ve ISOGG, Karafet ve diğerlerine karşı çıkıyor, tüm veriler aslında M81'e filogenetik olarak eşdeğer kabul edilmesi gerektiğini ima ediyor gibi görünüyor.[kaynak belirtilmeli ]
- E1b1b1b1b1 (M165) Underhill vd. (2000).
- E1b1b1b1b2 (L351) The E-M35 Filogeni Projesi.
- E1b1b1b1 (M81)
Ortalama Kuzey Afrikalı Faslı Berberiler, +% 80'de E3b3 frekansına sahiptir. Alvarez et al.(2009) Çalışma, E3b1b sıklığının 28/33 veya Berberilerde% 84,8 olduğunu göstermektedir. Marakeş. Geri kalan frekanslar 1/33 =% 3 E3a *, 1/33 =% 3 E3b *, 1/33 veya% 3 E3b1a ve 1/33 veya% 3 E3b1c.[31]
E1b1b (M81) Proto-Berberi işaretleyici E1b1b1b1a1 (M107) İndirgenmiş Proto-Berberi soy Mali.[41]
E-M78

En bazal ve nadir E-M78 * Paragrup Faslı Araplarda daha düşük frekanslarda bulunmuştur. Alt sınıf E-V65, yüksek seviyelerde bulunur. Mağrip uzak kuzey Afrika'nın bölgeleri. Cruciani vd. (2007) arasında yaklaşık% 20 düzeyinde Libya Arap soylar ve yaklaşık% 30 Faslı Araplar. Daha az yaygın gibi görünüyor Berberiler ancak yine de>% 10 seviyelerinde mevcuttur. Yazarlar bu soy için Kuzey Afrika kökenli olduğunu öne sürüyorlar. Avrupa'da, İtalya ve Yunanistan'da sadece birkaç kişi bulundu.
Capelli vd. (2009) Avrupa'daki beta kümesini inceledi. Güney İtalya'da küçük miktarlar buldular, ancak aynı zamanda Cantabria, Portekiz ve Galiçya'da da izler buldular, çalışmalarında Cantabria% 3,1 ile Avrupa'da en yüksek seviyeye sahipti (161 kişiden 5'i).
Diğer frekanslar E1b1b1a1c (E-V22) Cruciani ve ark. (2007) arasında Faslı Araplar (% 7,27, 55 kişi) ve Faslı Yahudiler (% 8, 50 kişi).
Fas Y-DNA Haplogrupları
| Nüfus | n | A / B | E-M33 | E-V38 | E-M35 * | E-M78 | E-M81 | E-M123 | G | J | R | Referans |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fas | 760 | 0.9 | 2.7 | 3.2 | 4.2 | 6.8 | 67.3 | 0.6 | 0.6 | 7.6 | 4.4 | Bekada vd. 2013[42] |
| Fas | 87 | — | — | 9.2 | — | 5.7 | 52.8 | — | — | 26.4 | — | Fadhlaoui-Zid vd. 2013[43] |
| Fas | 221 | — | 1.8 | 4.5 | 4 | 6.8 | 65 | — | — | 9 | 4 | Fregel vd. 2009[44] |
| Fas | 51 | 4 | 6 | — | 6 | 6 | 55 | — | — | 20 | 4 | Onofri vd. 2008[45] |
| Fas | 176 | — | — | 6.3 | 5.1 | 6.3 | 63.6 | — | — | 13.6 | 2.8 | Bosch ve diğerleri. 2001[46] |
| Araplar (Fas ) | 49 | — | — | — | — | 42.9 | 32.6 | — | — | 20.4 | — | Semino vd. 2004[47] |
| Araplar (Fas ) | 44 | — | — | 6.8 | 2.2 | 11.3 | 52.2 | — | — | 15.9 | 6.8 | Bosch ve diğerleri. 2001[46] |
| Araplar (Fas ) | 54 | — | — | — | — | 38.9 | 31.5 | — | — | — | — | Cruciani vd. 2004[48] |
| Berberiler (Fas ) | 64 | — | — | — | — | 10.9 | 68.7 | — | — | 6.3 | — | Semino vd. 2004[47] |
| Berberiler (Marakeş ) | 29 | — | — | — | 3.4 | 6.9 | 72.4 | — | — | — | — | Cruciani vd. 2004[49] |
| Berberiler (Orta Atlas ) | 69 | — | — | — | — | 10.1 | 71 | — | 4.3 | 5.8 | — | Cruciani vd. 2004[49] |
| Berberiler (Güney Fas ) | 40 | — | — | 2.5 | 7.5 | 12.5 | 65 | — | — | 10 | — | Bosch ve diğerleri. 2001[46] |
| Berberiler (Kuzey Merkez) | 63 | — | 3.1 | 9.5 | 7.9 | 1.5 | 65 | — | — | 11.1 | — | Bosch ve diğerleri. 2001[46] |
| Berberiler (Amizmiz) | 33 | 3 | — | 3 | 3 | 3 | 84.8 | 3 | — | — | — | Alvarez vd. 2009[50] |
| Berberiler (Asni) | 54 | — | — | — | 1.9 | 3.7 | 79.6 | — | — | 1.9 | 1.9 | Dugoujon vd. (2005)[51] |
| Berberiler (Bouhria) | 67 | — | — | — | — | 1.5 | 77.6 | — | 6 | 1.5 | 6 | Dugoujon vd. (2005)[51] |
| Berberiler (Kuzey Fas) | 43 | — | — | — | — | — | 79.1 | — | — | — | — | Ahmed Reguig vd. 2014[52] |
| Berberiler (Güney Fas) | 65 | — | — | — | — | — | 98.5 | — | — | — | — | Ahmed Reguig vd. 2014[52] |
| Berberiler (Orta Fas) | 187 | — | — | — | — | — | 89.8 | — | — | — | — | Ahmed Reguig vd. 2014[52] |
| Fas Sahra | 189 | 0.5 | 5.2 | 6.8 | — | — | 55.5 | 11.1 | — | 13.2 | 7.2 | Bekada vd. 2013[42] |
| Fas Sahra | 89 | — | 8.9 | 11.2 | — | — | 59.5 | — | — | 20.2 | — | Fregel vd. 2009[44] |
| Fas Sahra | 29 | — | 3.4 | 3.4 | — | — | 75.8 | — | — | 17.2 | — | Bosch ve diğerleri. 2001[46] |
| Faslı Yahudiler | 19 | — | — | — | 21.1 | — | — | — | 26.3 | 31.5 | 10.5 | Francalacci vd. 2008[53] |
J-P209

Haplogroup J-P209'un yaklaşık 31.700 yıl önce Güneybatı Asya (31.700 ± 12.800 yıl önce Semino 2004Haplogrup J-P209 en yüksek yoğunlukta Güneybatı'da bulunur. Arap Yarımadası. Bu bölgenin dışında, Haplogroup J-P209'un Kuzey Afrika: Cezayir (% 35'e kadar) (Semino 2004 ), Tunus (% 31'e kadar),[54] Fas (% 20'ye kadar) (Semino 2004 )Mısır (% 20'ye kadar) (Luis 2004 ).
Diğer haplogruplar

E-M34 SNP'yi kontrol etmeden E-M123 ile ilgili olarak Fas'ta küçük frekanslarda bulunur A Düşük bölgesel yüzdeler E-M123 rapor edildi Faslı Berberiler yaklaşık% 3. E-M123 E1b1b1b2a (ISOGG 2012) olarak da bilinir.
Avrasya haplogrupları gibi Haplogrup J ve Haplogrup R1 ayrıca çok düşük frekanslarda da gözlemlenmiştir. Cruciani ve diğerleri tarafından kapsamlı bir çalışma. (2004), Fas'taki popülasyonları analiz eden, Kuzey Afrika'daki Y kromozomal varyasyon modelinin (her ikisi de dahil) olduğu sonucuna varmıştır. J1 ve R1b haplogrupları) büyük ölçüde Neolitik kökenlidir, bu da dünyanın bu bölgesindeki Neolitik geçişe Berberi konuşan pastoralistlerin Cezayir Çölü'nden Doğu Fas'a demik yayılmasının eşlik ettiğini öne sürmektedir, ancak daha sonraki makaleler bu tarihin olabileceğini öne sürmüştü. Oranian'dan geçişle on bin yıl kadar uzun bir süre önce Capsian Kuzey Afrika'da kültür.[55][56]
G ve T haplogrupları Fas'ta nadiren bulunur. Fas,% 1 G olarak bulundu.[57]
Başka bir çalışmada Fas'taki 312 numunenin% 1'i G.[58]
Başka bir çalışma, yalnızca Fas'ın Azgour Vadisi'ndeki mezralardan örnekleri topladı ve burada 33 numuneden hiçbirinin tespit edilmedi G.[31] Bu mezralar, tipik olarak hissedildikleri için seçildi. Berberi kompozisyonda.
20 Faslı bir çalışma Yahudiler % 30 G idi.[31] Test edilen adamlar daha sonra görünüşe göre İsrail. Yahudi erkeklerle ilgili bir başka çalışmada, Faslı 83 Yahudi erkeğin% 19.3'ünün Haplogroup G'ye ait olduğu bulundu.[59] G'nin üzerindeki Fas örnekleri SNP G2a2b Haplogroup'ta Muhtemelen Pozitiftir, Avrupa'da MÖ 5000 ile 3000 yılları arasına tarihlenen neolitik insan kalıntılarında tespit edilmiştir. ayrıca, tüm erkek iskeletlerinin çoğu Avrupa Neolitik dönem şimdiye kadar bu haplogruba ait Y-DNA'yı mumyalanmış kalıntılar gibi vermiştir. Buz Adam Ötzi, National Geographic Topluluğu Haplogroup G'nin kökenlerini 30.000 yıl önce Orta Doğu'ya yerleştiriyor ve haplogrubu taşıyan insanların Neolitik Çağ'ın Afrika'ya ve ardından Avrupa'ya yayılmasında yer aldığını varsayıyor [60] Arap'ın yüzde ikisi Faslılar ve Asni Oasis'in Berberi Faslılarının% 0 ila% 8'i aynı şekilde G.[61]
Haplogrup T, merkezi Berberiler arasında bulunur. Asni Vahası % 1,9 ile Cezayir sınırlarına yakın ve Faslı Yahudiler % 4'te.
E1b1a, Fas'ta düşük frekanslarda bulunur, bu soylar, özellikle Büyük Çöl çevresinde, Sahra boyunca kölelik ticaretine bağlı olarak, bazı özel bölgelerde bulunur. Haratinler veya Gnawa Kuzey Orta Fas'ta Cezayir sınırlarına yakın, Sahrawiler, Faslı Araplar ve Güney Fas'ta bulunan Asni Oasis'in Berberileri arasında.[62]
En bazal ve nadir E1a * paragroup, Faslı Berberiler ve Sahrawis'ten elde edilen örneklerde daha düşük frekanslarda bulunmuştur. Paleolitik çağda E1b1b ile birlikte Yakın Doğu'dan Kuzey Afrika'ya Arka Avrasya Göçüne Bağlı olarak yaklaşık 45.000BC tarihli.[61]
Fas Popülasyonlarında Haplogrup dağılımları

Faslı Berberilerde bulunan Y-DNA haplogruplarının ana bileşenleri (E3b;% 94) Avrupalı ve komşu Kuzey Afrika ve Yakın Doğu popülasyonları ile paylaşılıyor. Haplogrupların küçük payları arasında Kuzey Batı Afrikalılar (E1a, A1a;% 1), Yakın Doğulular (J, G, T;% 2,4), Sahra Altı Afrikalılar (E3a;% 1,7) ve Avrupalılar ( R1b, I1;% 2) afinite.
Belirlenen başlıca yüzdelerin bazıları şunlardı:
- E1b1b: 56% (Aboukhalid 2010) - Tipik Afroasiatik konuşan popülasyonlar.
- J-P209: 20.4% (Semino 2004) - Arap yarımadasının tipik popülasyonları, Levant ve Kafkasya ılımlı bir dağılıma sahip Güneydoğu Avrupa, Kuzey Afrika, Afrika Boynuzu, Orta Asya ve Güney Asya.
- R1b:% 0,8 ila% 6,8 (Bosch 2001) - Tipik Batı Avrupalılar, biraz Batı Asya halklar Sudan Fulani ve Chadic -Orta Afrika'nın konuşan halkları ve bazı Orta Asya halkları (örneğin Başkurtlar, Türkmen ve Uygurlar ).[63]
- G:% 0,4 - Kafkasya'dan tipik insanlar ve daha az ölçüde Orta Doğu.
- A1a:% 0,5 - Tipik Haratin ile ilgili gruplar Kuzeybatı Afrika.
- E1a:% 0,5 - Aralarında bulunan nadir haplogrup Fas Berberiler, Sahrawis, Güney Avrupalılar ve Sahel'deki bazı Çadca konuşmacılar.
- E1b1a:% 1,7 - Tipik Nijer-Kongo konuşan popülasyonlar.
- T:% 0,4 - Batı Avrasya çevresinde yaygın olarak dağılmıştır.
- ben:% 0,4 - Kuzey ve Güneydoğu Avrupa'da zirveler ile günümüz Avrupalı popülasyonlarının çoğunda bulunabilir. Haplogrup I1 Y kromozomları da bazı popülasyonlar arasında bulunmuştur. Yakın Doğu Kafkasya, Kuzeydoğu Afrika ve Orta Sibirya.
Faslıların Berberi Genetik Kimliği

Berberilerin atası olan Fas'ın tarih öncesi nüfusları, daha geniş Paleo-Akdeniz halkları grubuyla ilişkiliydi. Afroasiatik aile muhtemelen Mezolitik dönemde, belki de Capsian kültürü.[64][65] DNA analizi, Berberi Faslı popülasyonları ile Faslılarınkiler arasında benzerlikler buldu. Sami halkı İskandinavya'nın yaklaşık 9.000 yıl öncesine ait bir bağlantı gösteriyor.[66]
MÖ 5000 civarında, Kuzey Afrika'nın nüfusu öncelikle Iberomaurusian ve Capsian kültürleri, daha yeni bir saldırı ile ilişkili Neolitik devrim.[67] Proto-Berberi kabileleri, Geç Tunçtan Erken Demir Çağı'na kadar bu tarih öncesi topluluklardan gelişti.[68]
Genetik Tarih Öncesi Genişlemeler
DNA kanıtı, Son Buzul Maksimum 25.000 ila 19.000 yıl önce, bir kilometreyi aşan kalınlıkta büyük buz tabakaları Kuzey Avrupa'nın büyük bölümünü kaplayarak bölgeyi insanlar için yaşanmaz hale getirdi. İnsan popülasyonlarının güneye, Akdeniz yakınlarındaki daha sıcak bölgelere çekildiğine inanılıyor. Bu dönemdeki sığınakların Iberia, Balkanlar ve İtalya. Biraz vardı gen akışı itibaren Fas içine Iberia.[69]
Buzulun maksimum seviyesinden sonra, Avrupa iklimi ısındığında, sığınakların Avrupa'nın yeniden doldurulduğu kaynak olduğu düşünülüyor. İber sığınağına getirilen Berberi soyları, daha sonra insanların kuzeye doğru genişlemesiyle tüm Avrupa'ya dağılmış olacaktı. Bu, Doğu Avrupa'da ve Rusya kadar kuzeyde, esas olarak Kuzeybatı Afrika ile tarih öncesi bağlantıları olduğu görülen genetik soyların varlığını açıklayabilir. Fas (görmek mtDNA ).[69] İber sığınaklarından gelen insan nüfusunun genişlemesinin de geri döndüğüne inanılıyor. Fas ve Kuzeybatı Afrika.[70]
Tarih öncesi dönemin sonuna kadar Neolitik
Dan değişim avlanma ve toplama sırasında tarıma Neolitik Devrim dünya tarihinde bir dönüm noktasıydı. Tarıma geçişi ilk gerçekleştiren toplumların, Kuzey Afrika ve Orta Doğu yaklaşık 10.000 BCE. Tarım, Orta Doğu'dan çiftçiler göç ederek Avrupa'ya tanıtıldı.[71] Göre demik difüzyon modeline göre, bu Orta Doğulu çiftçiler, "Afrika dışına" göçten bu yana Avrupa'da yaşayan yerel avcı-toplayıcı popülasyonların yerini aldı veya onlarla melezleşti.[72]
İlk Orta Doğulu çiftçilerin Kuzey Afrikalı esas olarak etkiler Capsian kültürü.[73] Orta Doğu'da bulunan bazı genetik soyların bu dönemde oraya geldiğine dair öneriler var.[74] Orta Doğu'daki ilk Tarım toplumlarının genellikle Natufian Kültürü Filistin'de MÖ 12.000 - MÖ 10.000 arasında var olan. Kuzeybatı Afrika'dan önemli bir göç, Ibero-Maurisians itibaren Fas karşısında Sina Natufian'ın oluşumundan önce meydana gelmiş gibi görünüyor.[75]
Fas'ta genetik süreklilik
2013 yılında, yapımcılarına ait iskeletler Epipaleolitik Iberomaurusian tarih öncesi sitelerinde kazılan kültür Taforalt ve Afalou, antik DNA için analiz edildi. Tüm örnekler, Epipaleolitik dönemden bu yana bu bölgeler arasında gen akışını gösteren Kuzey Afrika veya kuzey ve güney Akdeniz kıyılarıyla ilişkili anne sınıflarına aitti.[76] Eski Taforalt bireyleri Y-DNA haplogrupunu taşıdı E1b1b ve mtDNA haplogrupları U6, H, JT ve V Iberomaurusian döneminden kalan bölgede nüfus devamlılığına işaret ediyor.[77][78]
Faslı Arapların Genetik Berberi Miras sürekliliği
Kuzey Afrika'da Berberi ve Arap örnekleri arasında mevcut olan kültürel farklılaşma, AMOVA analizlerinde ve MDS ve PC analizlerinde gösterildiği gibi her iki grup arasındaki genetik farklılıkları yansıtmıyor gibi görünüyor. Eğer Araplar Kuzey Afrika'da çoğunlukla Orta Doğu Araplarının torunları vardı, Orta Doğu'da başka yerlere göre çok daha yaygın olan N, U1, U3, U7 ve HV gibi haplogrupların frekansları, Kuzey Afrika Araplarında Berberilerden daha büyük olmalı. Ancak bunun tersi gözlemleniyor: Bu haplogrupların toplamı Kuzey Afrika Araplarında% 5'e, Berberilerde% 10'a kadar çıkıyor.
Kuzey Afrika Arapları ve Berberiler arasında farklılaşma eksikliği, klasik belirteçler gibi diğer genetik belirteçler kullanılarak da gözlemlenmiştir (Bosch ve diğerleri, 1997); otozomal STR'ler (Bosch ve diğerleri 2000), Alu ekleme polimorfizmleri (Comas ve diğerleri 2000); ve Y-kromozom soyları Bu örüntü, bölgenin Araplaştırılmasının, Arap yayılmasının meydana geldiği bölgede yaşayan Berberi nüfuslarının demografik bir yer değiştirmesinden ziyade esas olarak kültürel bir süreç olduğunu göstermektedir.[79]
Fas Mitokondriyal mtDNA
Fas mitokondriyal havuzu esasen yapısında Berberidir ve "Batı Avrasya haplogruplarının genel olarak yüksek frekansı" ile karakterize edilir.Fas'taki mtDNA HV haplogrubunun ince karakterizasyonu ile ortaya çıkan İberya'dan Kuzey Afrika'ya son buzul maksimum genişlemesi ile temsil edilir. % 36 ila% 60, Sahra altı L soylarının bir şekilde daha düşük frekansı ve Kuzey Afrika haplogrupları U6 ve M1'in önemli (ancak farklı) varlığı ".[80] Ve Cherni ve ark. 2008 "İberya'da başlayan Son buzul sonrası maksimum genişleme, sadece Avrupa'nın değil, aynı zamanda Kuzey Afrika'nın da yeniden yerleşmesine yol açtı".[81]
Avrasya mtDNA (maternal) sekansları, Faslı Berberilerde% 96, Cezayirli Berberilerde% 82 ve Berberi olmayan Faslılarda% 78, Senegalli popülasyonda sadece% 4 sıklıkta tespit edildi. (Rando 1998)
Yakın zamana kadar, bazı gazeteler Fas'taki ana L haplogruplarının dağıtımının esas olarak Sahra-ötesi köle ticaretinden kaynaklandığını öne sürüyordu.[82] Ancak, Eylül 2010'da, Frigi tarafından Berber mtDNA'sı hakkında kapsamlı bir çalışma et al. L haplogruplarının çoğunun çok daha eski olduğu ve yaklaşık 20.000 yıl önce eski bir Afrika gen akışı tarafından tanıtıldığı sonucuna vardı.[83]
Faslı Kuzey ve Güney Berberileri, SSA mtDNA'sının yalnızca% 3 ila% 1'ine sahiptir, Gen havuzuna Sahra altı katkısında bu kuzey-güney gradyanı desteklenmektedir. Esteban vd.,[84] mtDNA soylarının geri kalanı için çoğunlukla Kafkas / Batı Avrasyalı iken, Faslı Araplar, L-mtDNA sekansları aracılığıyla yaklaşık% 21 ila% 36 arasında daha yüksek SSA maternal karışımına sahipken, L-mtDNA'nın en yüksek frekansları, Çevreleyen bölgedeki Faslı Araplara Bildirildi nın-nin El jadida % 36'da ve bu büyük ölçüde köle ticareti.[85]
L-mtDNA'nın frekansları (>% 1)
| Ülke | Etnik grup | Numara test edildi | Referans | % L-mtDNA |
| Fas | Faslı (Yahudiler) | 149 | Behar vd. (2008) | 1.34% |
| Fas | Faslı Kuzey (berberler) | 124 | Esteban vd. (2004) | 1% |
| Fas | Faslı (Araplar) | 81 | Harich vd. (2010) | 36% |
| Fas | Faslı Araplar | 56 | Turchi vd. (2009) | 25.00% |
| Fas | Faslı Güney (Berberiler) | 64 | Turchi vd. (2009) | 26.00% |
Faslıların Güney Avrupa üzerindeki genetik etkisi

Trombetta vd. (2011), V257'nin kardeş sınıfıyla bir paralellik gösterdiğini hissetti E-V68 Her iki grubun da Kuzey Batı Afrika'dan Akdeniz üzerinden güneybatı Avrupa'ya göç ettiğine dair işaretler göstermesi gibi. Örneklerinde Faslı Marakeş Berberi, Korsikalı, Sardunyalı, güney İspanyol ve Kantabria'dan E-V257 olan 6 "E-V257 *" birey buldular.
E-M35'te iki haplogrup, E-V68 ve E-V257 arasında çarpıcı paralellikler var. Her ikisi de çoğunlukla Fas'ta (sırasıyla E-M78 ve E-M81) Kuzey Batı Afrika'da sıklıkla gözlemlenen bir soy ve çoğunlukla Güney Avrupa'da bulunan bir grup farklılaşmamış kromozom içerir. Kuzey Afrika'da E-V68 * ve E-V257 * 'nin E-M35 taşıyıcılarının genişlemesi, Fas ve güney Avrupa arasında denizcilikte yayılmayı daha makul bir hipotez haline getiriyor.
Semino'dan yapılan bir araştırma (2004'te yayınlandı) şunu gösterdi: Y kromozomu haplotip E1b1b1b (E-M81), özeldir Fas nüfusu ve Avrupa'da neredeyse yok Iberia (ispanya ve Portekiz ) ve Sicilya.
Haplogroup E1b1b1b'nin Güney İspanya'da% 10'luk frekansları geçtiğine göre İber yarımadasının Y kromozom analizi. Çalışma, hem tarihi hem de tarih öncesi çağlarda Kuzey Afrika ve Orta Doğu'dan çok sınırlı bir etkiye işaret ediyor.[86] Mikrosatellit varyasyonunun yokluğu, Fas'tan İslami yayılma döneminde Akdeniz'deki tarihsel değişimlerle, yani Berberi popülasyonlarıyla tutarlı bir şekilde çok yakın zamanda geldiğine işaret ediyor. Sınırlı bir çalışma Portekiz Y kromozom soyları ile ilgili olarak, "mtDNA ve Y verileri, o bölgedeki Berberi varlığının MS 711'deki Mağribi yayılmasından önceye dayandığını göstermektedir. ... Verilerimiz, Sahra altı göçmenlerin aksine erkek Berberilerin oluştuğunu göstermektedir. ülkede uzun ömürlü ve sürekli bir topluluk ".[87]
Karakteristik bir Fas haplotipi olan Haplotip V (p49 / TaqI), İber yarımadasında da bulunabilir ve azalan Kuzey-Güney frekansı, Fas'tan İberya'ya doğru aynı zamanda tutarlı bir gen akışı oluşturur. Mağribi yarımadadaki varlığı.[88] Haplotip V'in Kuzey-Güney sıklığı, güney Portekiz'de% 30'a yakın frekanslardan güney Fransa'da yaklaşık% 10'a kadar değişen Akdeniz bölgesinde gözlemlenecektir. Benzer şekilde, İtalya'da en yüksek sıklık güneydeki Sicilya adasında görülmektedir (% 28).[89][90]
81 popülasyondan 6.501 ilgisiz Y kromozom örneğini kullanan geniş kapsamlı bir çalışma (2007'de yayınlandı) şunu buldu: "Hem bu E-M78 alt haplogrupları (E-V12, E-V22, E-V65) hem de E-M81 göz önüne alındığında haplogroup, Fas soylarının tüm erkek gen havuzuna katkısı Iberia (Pasiegos hariç), kıta İtalya ve Sicilya sırasıyla% 5,6,% 3,6 ve% 6,6 olarak tahmin edilebilir. "[90]
Hakkında bir çalışma Sicilya Gaetano tarafından et al. 2008, "Kuzeybatı Afrika Faslı popülasyonlarında yaygın olarak yayılan Hg E3b1b-M81'in Sicilya gen havuzuna% 6 oranında katkıda bulunduğu tahmin edilmektedir."[91]
"Çalışma, din değiştirmelerin ve farklı soydan insanlar arasındaki sonraki evliliklerin hem İspanya'da, özellikle Balear Adaları'nda hem de Portekiz'de modern nüfus üzerinde önemli bir etkiye sahip olduğunu gösteriyor",[92]
Avrupa'da, E-M81 her yerde bulunur, ancak çoğunlukla Iber Yarımadası, Avrupa'nın geri kalanının aksine[Not 1] E-M78'den daha yaygındır ve ortalama frekansı% 5 civarındadır.[57][93][94][95][96] Bu sınıfın şu ana kadar Avrupa'da bulunan en yüksek frekansları, Pasiegos itibaren Cantabria,% 18 (8/45) arasında değişen[96] % 41'e (23/56).[49] İspanyolcada da ortalama% 8,28 (54/652) sıklığı rapor edilmiştir. Kanarya Adaları en büyük üç adada% 10'un üzerinde sıklık ile Tenerife (10.68%), Gran Canaria (% 11,54) ve Fuerteventura (13.33%).[97]
Latin Amerika üzerindeki genetik etkiler
Latin Amerika'nın İspanyol ve Portekiz kolonizasyonunun bir sonucu olarak, E-M81 aynı zamanda Latin Amerika[98][99][100] ve ABD’deki Latin Amerikalı erkekler arasında.[101]
Diğer bölgeler
Diğer ülkelerde, Fas Berberi haplogrupları şurada bulunabilir: Mısır, Fransa, Sudan, Somali, Ürdün, Lübnan, Suudi Arabistan ve arasında Sefarad Yahudileri.[94]
Referanslar
- ^ . Ancak bu çalışma aynı zamanda Mayorka, Minorka ve İbiza adalarından 153 kişiyi ve İber Yarımadası'nda olmayan Gaskonya'dan 24 kişiyi kapsamaktadır. Bu 177 kişi olmadan, İber Yarımadası için gerçek ortalama% 4,9 (47/963), tabloya bakın Arşivlendi 2013-07-30 Wayback Makinesi.
- ^ Saha Projeleri - Jebel Irhoud [https://web.archive.org/web/20170112002657/http://www.eva.mpg.de/evolution/files/irhoud.htm Arşivlendi 2017-01-12 de Wayback Makinesi. İnsan Evrimi Bölümü. Max Planck Evrimsel Antropoloji Enstitüsü
- ^ Kızamıkçık, D. (1984). "Mağrip'te Çevrecilik ve Pi Paleolitik ekonomiler (MÖ 20.000 ila 5.000 M.Ö.)". J.D. Clark & S.A. Brandt (ed.). From hunters to farmers the causes and consequences of food production in Africa. Berkeley: California Üniversitesi Yayınları. pp. 41–56. ISBN 978-0520045743.
- ^ The Megalithic Portal; Megalith Map. "C. Michael Hogan, Mogador: Promontory Fort, The Megalithic Portal, ed. Andy Burnham". Megalithic.co.uk. Alındı 2010-06-02.
- ^ Moscati, Sabatino (2001) The Phoenicians, Tauris, ISBN 1-85043-533-2
- ^ "Bocchus I". Alındı 27 Eylül 2010.
- ^ Ramirez-Faria, Carlos (2007-01-01). Concise Encyclopaedia of World History. ISBN 978-81-269-0775-5.
- ^ "Almoravides". Universalis Encyclopedia.
- ^ "Marīnid dynasty". Encyclopædia Britannica.
- ^ "The Maghrib under the Almoravids and the Almohads". Britannica.com. Alındı 2011-08-01.
- ^ "Morocco – History". Britannica.com. Alındı 2011-08-01.
- ^ Allo Isichei, Elizabeth (1997). A history of African societies to 1870. Cambridge University Press. s. 264. ISBN 0-521-45599-5
- ^ "Morocco (Page 8 of 9) ". Microsoft Encarta Online Encyclopedia 2009. 2009-11-01.
- ^ "Joint Statement by the United States of America and the Kingdom of Morocco". 2013-11-22.
- ^ International Business Publications, USA (2004). Fas Dış Politikası ve Devlet Rehberi. ISBN 9780739760000.
- ^ "Defense Department News".
- ^ Kozaryn, Linda D. "Cohen ABD-Fas Bağlarını Yeniledi". ABD Savunma Bakanlığı. Alındı 2009-03-12.
- ^ Roberts, Priscilla H. ve Richard S. Roberts, Thomas Barclay (1728–1793): Consul in France, Diplomat in Barbary, Lehigh University Press, 2008, pp. 206–223 ISBN 093422398X.
- ^ "Amerikan Diplomasisinin Kilometre Taşları, İlginç Tarihi Notlar ve Dışişleri Bakanlığı Tarihi". ABD Dışişleri Bakanlığı. Alındı 2007-12-17.
- ^ Coudray, C.; Olivieri, A.; Achilli, A.; Pala, M.; Melhaoui, M.; Cherkaoui, M.; El-Chennawi, F.; Kossmann, M.; Torroni, A.; Dugoujon, J. M. (2009). "The Complex and Diversified Mitochondrial Gene Pool of Berber Populations". İnsan Genetiği Yıllıkları. 73 (2): 196–214. doi:10.1111/j.1469-1809.2008.00493.x. PMID 19053990.
- ^ Henn, B. M.; Botigué, L. R.; Gravel, S.; Wang, W .; Brisbin, A.; Byrnes, J. K.; Fadhlaoui-Zid, K.; Zalloua, P. A.; Moreno-Estrada, A. (2012). Schierup, Mikkel H (ed.). "Genomic Ancestry of North Africans Supports Back-to-Africa Migrations". PLOS Genetiği. 8 (1): e1002397. doi:10.1371/journal.pgen.1002397. PMC 3257290. PMID 22253600.
- ^ Ballais, Jean-Louis (2000) "Chapter 7: Conquests and land degradation in the eastern Maghreb" İçinde Barker, Graeme and Gilbertson, David (2000) The Archaeology of Drylands: Living at the Margin Routledge, London, Volume 1, Part III – Sahara and Sahel, pp. 125–136, ISBN 978-0-415-23001-8
- ^ Gomez-Casado, E.; Del Moral, P.; Martinez-Laso, J.; Garcia-Gomez, A.; Allende, L.; Silvera-Redondo, C.; Longas, J.; Gonzalez-Hevilla, M.; Kandil, M.; et al. (2000). "HLA genes in Arabic-speaking Moroccans: Close relatedness to Berbers and Iberians". Doku Antijenleri. 55 (3): 239–249. doi:10.1034/j.1399-0039.2000.550307.x. PMID 10777099.
- ^ Carr, Matthew (2009). Blood and faith: the purging of Muslim Spain. Yeni Basın. s. 290. ISBN 978-1-59558-361-1.
- ^ Bosch, E .; Calafell, F.; Pérez-Lezaun, A.; Clarimón, J.; Comas, D.; Mateu, E.; Martínez-Arias, R.; Morera, B.; Brakez, Z. (2000). "Genetic structure of north-west Africa revealed by STR analysis". European Journal of Human Genetics. 8 (5): 360–366. doi:10.1038/sj.ejhg.5200464. PMID 10854096.
- ^ Harich, N.; Esteban, E.; Chafik, A.; López-Alomar, A.; Vona, G.; Moral, P. (2002). "Classical polymorphisms in Berbers from Moyen Atlas (Morocco): Genetics, geography, and historical evidence in the Mediterranean peoples". İnsan Biyolojisi Yıllıkları. 29 (5): 473–487. doi:10.1080/03014460110104393. PMID 12396367.
- ^ Arredi, B.; Poloni, E.; Paracchini, S.; Zerjal, T.; Fathallah, D.; Makrelouf, M.; Pascali, V.; Novelletto, A.; Tyler-Smith, C. (2004). "A Predominantly Neolithic Origin for Y-Chromosomal DNA Variation in North Africa". The American Journal of Human Genetics. 75 (2): 338–345. doi:10.1086/423147. PMC 1216069. PMID 15202071.
- ^ Henn et al. (2008)
- ^ Hassan et al. (2008)
- ^ Cruciani, F.; La Fratta, R.; Santolamazza, P.; Sellitto, D.; Pascone, R.; Moral, P.; Watson, E.; Guida, V.; Colomb, E. B. (2004). "Haplogroup E3b (E-M215) Y Kromozomlarının Filocoğrafik Analizi, Afrika İçinde ve Dışında Çoklu Göç Olaylarını Gösteriyor". The American Journal of Human Genetics. 74 (5): 1014–1022. doi:10.1086/386294. PMC 1181964. PMID 15042509.
- ^ a b Arredi et al. (2004)
- ^ a b c d Alvarez, L.; Santos, C.; Montiel, R.; Caeiro, B.; Baali, A.; Dugoujon, J. M.; Aluja, M. P. (2009). "Y-chromosome variation in South Iberia: Insights into the North African contribution". Amerikan İnsan Biyolojisi Dergisi. 21 (3): 407–409. doi:10.1002 / ajhb.20888. PMID 19213004.
- ^ Ehret et al. (2004)
- ^ Keita & Boyce (2005)
- ^ "E-M81 YTree". yfull.com. Alındı 2016-07-10.
- ^ "E-M215 YTree". yfull.com. Alındı 2016-07-10.
- ^ Alvarez et al. 2009
- ^ Kujanová, Martina; Pereira, Luísa; Fernandes, Verónica; Pereira, Joana B .; Cerný, Viktor (October 2009). "Mısır Batı Çölü'nün küçük bir vahasında yakın doğu neolitik genetik girdisi". Amerikan Fiziksel Antropoloji Dergisi. 140 (2): 336–346. doi:10.1002 / ajpa.21078. ISSN 1096-8644. PMID 19425100.
- ^ ISOGG (2008)
- ^ Karafet, T. M .; Mendez, F.L .; Meilerman, M. B .; Underhill, P. A .; Zegura, S. L .; Çekiç, M.F. (2008). "Yeni ikili polimorfizmler, insan Y kromozom haplogrup ağacını yeniden şekillendirir ve çözünürlüğünü artırır". Genom Araştırması. 18 (5): 830–838. doi:10.1101 / gr.7172008. PMC 2336805. PMID 18385274.
- ^ Consortium, T. Y. C. (2002). "A Nomenclature System for the Tree of Human Y-Chromosomal Binary Haplogroups". Genom Araştırması. 12 (2): 339–348. doi:10.1101 / gr.217602. PMC 155271. PMID 11827954.
- ^ "The Genetic Atlas - E1b1b Meditid mutative history". thegeneticatlas.com. Alındı 2015-10-01.
- ^ a b Bekada, Asmahan; Fregel, Rosa; Cabrera, Vicente M .; Larruga, José M .; Pestano, José; Benhamamouch, Soraya; González, Ana M. (2013-02-19). "Cezayir Mitokondriyal DNA'sını ve Y-Kromozom Profillerini Kuzey Afrika Manzarasına Tanıtmak". PLOS ONE. 8 (2): e56775. Bibcode:2013PLoSO ... 856775B. doi:10.1371 / journal.pone.0056775. ISSN 1932-6203. PMC 3576335. PMID 23431392.
- ^ Fadhlaoui-Zid et al. 2013, Genome-Wide and Paternal Diversity Reveal a Recent Origin of Human Populations in North Africa
- ^ a b Fregel vd. 2009, Demographic history of Canary Islands male gene-pool: replacement of native lineages by European.
- ^ Onofri et al. 2008, Y-chromosome markers distribution in Northern Africa: High-resolution SNP and STR analysis in Tunisia and Morocco populations
- ^ a b c d e Bosch et al. 2001, High-resolution analysis of humanY-chromosome variation shows a sharp discontinuity and limited gene flow between Northwestern Africa and the IberianPeninsula.
- ^ a b Semino, O.; Magri, C .; Benuzzi, G.; Lin, A. A .; Al-Zahery, N.; Battaglia, V.; MacCioni, L.; Triantaphyllidis, C .; Shen, P. (2004). "Y-Kromozom Haplogrupları E ve J'nin Kökeni, Difüzyonu ve Farklılaşması: Avrupa'nın Neolitikleşmesi ve Daha Sonra Akdeniz Bölgesi'ndeki Göç Olayları Üzerine Çıkarımlar". The American Journal of Human Genetics. 74 (5): 1023–34. doi:10.1086/386295. PMC 1181965. PMID 15069642.
- ^ Cruciani et al. 2004, Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out Of Africa.
- ^ a b c Cruciani, F.; La Fratta, R.; Santolamazza, P.; Sellitto, D.; Pascone, R.; Moral, P.; Watson, E.; Guida, V.; Colomb, E. B. (2004). "Haplogroup E3b (E-M215) Y Kromozomlarının Filocoğrafik Analizi, Afrika İçinde ve Dışında Çoklu Göç Olaylarını Gösteriyor". The American Journal of Human Genetics. 74 (5): 1014–1022. doi:10.1086/386294. PMC 1181964. PMID 15042509.
- ^ Alvarez, Luis; Santos, Cristina; Montiel, Rafael; Caeiro, Blazquez; Baali, Abdellatif; Dugoujona, Jean-Michel; Dugoujon, Jean-Michel; Aluja, Maria Pilar (2009-06-01). "Güney İberya'daki Y kromozomu varyasyonu: Kuzey Afrika'nın katkısına ilişkin bilgiler". Amerikan İnsan Biyolojisi Dergisi. 21 (3): 407–409. doi:10.1002 / ajhb.20888. ISSN 1520-6300. PMID 19213004.
- ^ a b "Berberiler: Dilsel ve genetik çeşitlilik" (PDF). ddl.ish-lyon.cnrs.fr. Arşivlenen orijinal (PDF) 25 Mart 2012 tarihinde. Alındı 22 Mart 2015.
- ^ a b c Ahmed, Reguig; Nourdin, Harich; Abdelhamid, Barakat; Hassan, Rouba (2014-01-01). "Phylogeography of E1b1b1b-M81 Haplogroup and Analysis of its Subclades in Morocco". Human Biology Open Access Pre-Prints. 86 (2).
- ^ Francalacci et al. 2008, History and geography of human Y-chromosome in Europe: a SNP perspective
- ^ 31% is based on Combined Data
- Semino 2004: 30%
- Arredi 2004: 32%
- ^ Semino, O.; Magri, C .; Benuzzi, G.; Lin, A. A .; Al-Zahery, N.; Battaglia, V.; MacCioni, L.; Triantaphyllidis, C .; Shen, P. (2004). "Y-Kromozom Haplogrupları E ve J'nin Kökeni, Difüzyonu ve Farklılaşması: Avrupa'nın Neolitikleşmesi ve Daha Sonra Akdeniz Bölgesi'ndeki Göç Olayları Üzerine Çıkarımlar". The American Journal of Human Genetics. 74 (5): 1023–34. doi:10.1086/386295. PMC 1181965. PMID 15069642.
- ^ Myles, S.; Bouzekri, N.; Haverfield, E.; Cherkaoui, M.; Dugoujon, J. M.; Ward, R. (2005). "Genetic evidence in support of a shared Eurasian-North African dairying origin". İnsan Genetiği. 117 (1): 34–42. doi:10.1007/s00439-005-1266-3. PMID 15806398.
- ^ a b Adams, S. M .; Bosch, E .; Balaresque, P. L.; Ballereau, S. P. J.; Lee, A. C.; Arroyo, E.; López-Parra, A. M.; Aler, M.; Grifo, M. S. G. (2008). "The Genetic Legacy of Religious Diversity and Intolerance: Paternal Lineages of Christians, Jews, and Muslims in the Iberian Peninsula". The American Journal of Human Genetics. 83 (6): 725–736. doi:10.1016 / j.ajhg.2008.11.007. PMC 2668061. PMID 19061982.
- ^ Zalloua, P. A.; Platt, D. E.; El Sibai, M.; Khalife, J.; Makhoul, N.; Haber, M .; Xue, Y.; Izaabel, H.; Bosch, E. (2008). "Tarihsel Genişlemelerin Genetik İzlerini Tanımlamak: Akdeniz'deki Fenike Ayak İzleri". The American Journal of Human Genetics. 83 (5): 633–42. doi:10.1016 / j.ajhg.2008.10.012. PMC 2668035. PMID 18976729.
- ^ Behar, D. M.; Yunusbayev, B.; Metspalu, M.; Metspalu, E.; Rosset, S.; Parik, J. R.; Rootsi, S.; Chaubey, G.; Kutuev, I. (2010). "The genome-wide structure of the Jewish people". Doğa. 466 (7303): 238–242. Bibcode:2010Natur.466..238B. doi:10.1038/nature09103. PMID 20531471.
- ^ Alonso, S.; Flores, C.; Cabrera, V.; Alonso, A.; Martín, P.; Albarrán, C.; Izagirre, N.; De La Rúa, C. N.; García, O. (2005). "The place of the Basques in the European Y-chromosome diversity landscape". European Journal of Human Genetics. 13 (12): 1293–1302. doi:10.1038/sj.ejhg.5201482. PMID 16094307.
- ^ a b Cruciani, F.; Santolamazza, P.; Shen, P.; MacAulay, V.; Moral, P.; Olckers, A.; Modiano, D.; Holmes, S.; Destro-Bisol, G. (2002). "A Back Migration from Asia to Sub-Saharan Africa is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes". The American Journal of Human Genetics. 70 (5): 1197–1214. doi:10.1086/340257. PMC 447595. PMID 11910562.
- ^ Bosch, E .; Calafell, F.; Comas, D.; Oefner, P. J.; Underhill, P. A .; Bertranpetit, J. (2001). "High-Resolution Analysis of Human Y-Chromosome Variation Shows a Sharp Discontinuity and Limited Gene Flow between Northwestern Africa and the Iberian Peninsula". The American Journal of Human Genetics. 68 (4): 1019–29. doi:10.1086/319521. PMC 1275654. PMID 11254456.
- ^ Y Haplogroups of the World Online Baskı
- ^ Abdallah Laroui, The History of the Maghrib (Paris 1970; Princeton 1977) at 17, 60 (re S.W.Asians, referencing the earlier work of Gsell).
- ^ Camps, Gabriel (1996), Les Berbères, Edisud, pp. 11–14, 65
- ^ Achilli, A.; Rengo, C.; Battaglia, V.; Pala, M.; Olivieri, A.; Fornarino, S.; Magri, C .; Scozzari, R.; Babudri, N. (2005). "Saami and Berbers—An Unexpected Mitochondrial DNA Link". The American Journal of Human Genetics. 76 (5): 883–886. doi:10.1086/430073. PMC 1199377. PMID 15791543.
- ^ J. Desanges, "The proto-Berbers" 236-245, at 237, in General History of Africa, v.II Ancient Civilizations of Africa (UNESCO 1990).
- ^ Mário Curtis Giordani, História da África. Anterior aos descobrimentos (Petrópolis, Brasil: Editora Vozes 1985) at 42-43, 77-78. Giordani references Bousquet, Les Berbères (Paris 1961).
- ^ a b Malyarchuk, B. A .; Derenko, M.; Perkova, M.; Grzybowski, T .; Vanecek, T.; Lazur, J. (2008). "Reconstructing the phylogeny of African mitochondrial DNA lineages in Slavs". European Journal of Human Genetics. 16 (9): 1091–1096. doi:10.1038/ejhg.2008.70. PMID 18398433.
- ^ Cherni, L.; Fernandes, V. N.; Pereira, J. B.; Costa, M. D.; Goios, A.; Frigi, S.; Yacoubi-Loueslati, B.; Amor, M. B .; Slama, A. (2009). "Post-last glacial maximum expansion from Iberia to North Africa revealed by fine characterization of mtDNA H haplogroup in Tunisia". Amerikan Fiziksel Antropoloji Dergisi. 139 (2): 253–260. doi:10.1002/ajpa.20979. PMID 19090581.
- ^ Brace, C. L. (2006). "The questionable contribution of the Neolithic and the Bronze Age to European craniofacial form". Ulusal Bilimler Akademisi Bildiriler Kitabı. 103 (1): 242–247. Bibcode:2006PNAS..103..242B. doi:10.1073/pnas.0509801102. PMC 1325007. PMID 16371462.
- ^ Cavalli-Sforza, L.; Menozzi, P.; Piazza, A. (1993). "Demic expansions and human evolution". Bilim. 259 (5095): 639–646. Bibcode:1993Sci...259..639C. doi:10.1126/science.8430313. PMID 8430313.
- ^ Bar-Yosef O (1987) Pleistocene connections between Africa and Southwest Asia: an archaeological perspective. Afrika Arkeolojik İncelemesi; Chapter 5, pg 29-38.
- ^ Underhill, P. A .; Kivisild, T. (2007). "Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations". Annual Review of Genetics. 41: 539–564. doi:10.1146 / annurev.genet.41.110306.130407. PMID 18076332. S2CID 24904955.
- ^ Delson, Eric; Tattersall, Ian; Couvering, John Van; Brooks, Alison S. (2004-11-23). Encyclopedia of Human Evolution and Prehistory: Second Edition. Routledge. s. 97. ISBN 9781135582272.
- ^ Kefi R, Bouzaid E, Stevanovitch A, Beraud-Colomb E. "MITOCHONDRIAL DNA AND PHYLOGENETIC ANALYSIS OF PREHISTORIC NORTH AFRICAN POPULATIONS" (PDF). ISABS. Arşivlenen orijinal (PDF) 11 Mart 2016 tarihinde. Alındı 17 Ocak 2016.
- ^ Bernard Secher; Rosa Fregel; José M Larruga; Vicente M Cabrera; Phillip Endicott; José J Pestano; Ana M González (2014). "Kuzey Afrika mitokondriyal DNA haplogrup U6 geninin tarihi Afrika, Avrasya ve Amerika kıtalarına akıyor". BMC Evrimsel Biyoloji. 14: 109. doi:10.1186/1471-2148-14-109. PMC 4062890. PMID 24885141.
- ^ Loosdrecht, Marieke van de; Bouzouggar, Abdeljalil; Humphrey, Louise; Posth, Cosimo; Barton, Nick; Aximu-Petri, Ayinuer; Nickel, Birgit; Nagel, Sarah; Talbi, El Hassan (2018-03-15). "Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations". Bilim. 360 (6388): 548–552. Bibcode:2018Sci...360..548V. doi:10.1126/science.aar8380. ISSN 0036-8075. PMID 29545507.
- ^ Fadhlaoui-Zid, K.; Plaza, S.; Calafell, F.; Ben Amor, M.; Comas, D.; Bennamar, A.; Gaaied, E. (2004). "Mitochondrial DNA Heterogeneity in Tunisian Berbers". İnsan Genetiği Yıllıkları. 68 (3): 222–33. doi:10.1046/j.1529-8817.2004.00096.x. PMID 15180702. S2CID 6407058.
- ^ Coudray, C.; Olivieri, A.; Achilli, A.; Pala, M.; Melhaoui, M.; Cherkaoui, M.; El-Chennawi, F.; Kossmann, M.; Torroni, A.; Dugoujon, J. M. (2009). "The Complex and Diversified Mitochondrial Gene Pool of Berber Populations". İnsan Genetiği Yıllıkları. 73 (2): 196–214. doi:10.1111/j.1469-1809.2008.00493.x. PMID 19053990.
- ^ Cherni, L.; Fernandes, V. N.; Pereira, J. B.; Costa, M. D.; Goios, A.; Frigi, S.; Yacoubi-Loueslati, B.; Amor, M. B .; Slama, A. (2009). "Post-last glacial maximum expansion from Iberia to North Africa revealed by fine characterization of mtDNA H haplogroup in Tunisia". Amerikan Fiziksel Antropoloji Dergisi. 139 (2): 253–260. doi:10.1002/ajpa.20979. PMID 19090581.
- ^ Harich, N.; Costa, M. D.; Fernandes, V. N.; Kandil, M.; Pereira, J. B.; Silva, N. M.; Pereira, L. S. (2010). "The trans-Saharan slave trade - clues from interpolation analyses and high-resolution characterization of mitochondrial DNA lineages". BMC Evrimsel Biyoloji. 10: 138. doi:10.1186/1471-2148-10-138. PMC 2875235. PMID 20459715.
- ^ Frigi, S.; Cherni, L.; Fadhlaoui-Zid, K.; Benammar-Elgaaied, A. (August 2010). "Ancient Local Evolution of African mtDNA Haplogroups in Tunisian Berber Populations". İnsan biyolojisi. 82 (4): 367–384. doi:10.3378/027.082.0402. PMID 21082907.
- ^ Esteban, E.; González-Pérez, E.; Harich, N.; López-Alomar, A.; Via, M.; Luna, F.; Moral, P. (2004). "Genetic relationships among Berbers and South Spaniards based on CD4 microsatellite/Alu haplotypes". İnsan Biyolojisi Yıllıkları. 31 (2): 202–212. doi:10.1080/03014460310001652275. PMID 15204363.
- ^ Harich et al 2010
- ^ "Reduced Genetic Structure for Iberian Peninsula: implications for population demography. (2004)" (PDF). Arşivlenen orijinal (PDF) 2008-04-06 tarihinde. Alındı 2011-08-01.
- ^ Goncalves, R.; Freitas, A.; Branco, M.; Rosa, A.; Fernandes, A. T.; Zhivotovsky, L. A.; Underhill, P. A .; Kivisild, T.; Brehm, A. (2005). "Y-chromosome Lineages from Portugal, Madeira and Acores Record Elements of Sephardim and Berber Ancestry". İnsan Genetiği Yıllıkları. 69 (4): 443–54. doi:10.1111 / j.1529-8817.2005.00161.x. PMID 15996172. S2CID 3229760.
- ^ Lucotte, G.; Gerard, N.; Mercier, G. (2001). "North African Genes in Iberia Studied by Y-Chromosome DNA Haplotype 5". İnsan biyolojisi. 73 (5): 763–769. doi:10.1353/hub.2001.0066. PMID 11758696.
- ^ Gérard, N.; Berriche, S.; Aouizérate, A.; Diéterlen, F.; Lucotte, G. R. (2006). "North African Berber and Arab Influences in the Western Mediterranean Revealed by Y-Chromosome DNA Haplotypes". İnsan biyolojisi. 78 (3): 307–316. doi:10.1353/hub.2006.0045. PMID 17216803.
- ^ a b Cruciani, F.; La Fratta, R.; Trombetta, B.; Santolamazza, P.; Sellitto, D.; Colomb, E. B.; Dugoujon, J. -M.; Crivellaro, F .; Benincasa, T. (2007). "Kuzey / Doğu Afrika ve Batı Avrasya'da Geçmiş İnsan Erkek Hareketlerinin İzini Sürmek: Y-Kromozomal Haplogrupları E-M78 ve J-M12'den Yeni İpuçları". Moleküler Biyoloji ve Evrim. 24 (6): 1300–1311. doi:10.1093 / molbev / msm049. PMID 17351267.
- ^ Di Gaetano, C .; Cerutti, N .; Crobu, F .; Robino, C .; Inturri, S .; Gino, S.; Guarrera, S.; Underhill, P. A .; King, R. J .; et al. (2008). "Sicilya'ya farklı Yunan ve kuzey Afrika göçleri, Y kromozomundan elde edilen genetik kanıtlarla destekleniyor". European Journal of Human Genetics. 17 (1): 91–99. doi:10.1038 / ejhg.2008.120. PMC 2985948. PMID 18685561. "The co-occurrence of the Berber E3b1b-M81 (2.12%) and of the Mid-Eastern J1-M267 (3.81%) Hgs together with the presence of E3b1a1-V12, E3b1a3-V22, E3b1a4-V65 (5.5%) support the hypothesis of intrusion of Moroccan genes. (...) These Hgs are common in northern Africa and are observed only in Mediterranean Europe and together the presence of the E3b1b-M81 highlights the genetic relationships between the Moroccan Almohad Empire and Sicily. (...) Hg E3b1b-M81 network cluster confirms the genetic affinity between Sicily and The Moroccan Empires."
- ^ The religious conversions of Jews and Muslims have had a profound impact on the population of the Iberian Peninsula, Elena Bosch, 2008 Arşivlendi May 21, 2009, at the Wayback Makinesi
- ^ Capelli, C .; Onofri, V.; Brisighelli, F .; Boschi, I.; Scarnicci, F.; Masullo, M.; Ferri, G.; Tofanelli, S.; Tagliabracci, A. (2009). "Moors and Saracens in Europe: Estimating the medieval North African male legacy in southern Europe". European Journal of Human Genetics. 17 (6): 848–852. doi:10.1038/ejhg.2008.258. PMC 2947089. PMID 19156170.
- ^ a b Flores, C.; Maca-Meyer, N.; Larruga, J. M.; Cabrera, V. M.; Karadsheh, N.; Gonzalez, A. M. (2005). "Isolates in a corridor of migrations: A high-resolution analysis of Y-chromosome variation in Jordan". İnsan Genetiği Dergisi. 50 (9): 435–441. doi:10.1007 / s10038-005-0274-4. PMID 16142507.
- ^ Beleza, S.; Gusmao, L.; Lopes, A.; Alves, C.; Gomes, I.; Giouzeli, M.; Calafell, F.; Carracedo, A.; Amorim, A. (2006). "Micro-Phylogeographic and Demographic History of Portuguese Male Lineages". İnsan Genetiği Yıllıkları. 70 (2): 181–194. doi:10.1111 / j.1529-8817.2005.00221.x. PMID 16626329. S2CID 4652154.
- ^ a b Maca-Meyer, N.; Sanchez-Velasco, P.; Flores, C.; Larruga, J. -M.; Gonzalez, A. -M.; Oterino, A.; Leyva-Cobian, F. (2003). "Y Chromosome and Mitochondrial DNA Characterization of Pasiegos, a Human Isolate from Cantabria (Spain)". İnsan Genetiği Yıllıkları. 67 (4): 329–39. CiteSeerX 10.1.1.584.4253. doi:10.1046 / j.1469-1809.2003.00045.x. PMID 12914567.
- ^ Fregel, R.; Gomes, V. N.; Gusmão, L.; González, A. M.; Cabrera, V. M.; Amorim, A. N.; Larruga, J. M. (2009). "Demographic history of Canary Islands male gene-pool: Replacement of native lineages by European". BMC Evrimsel Biyoloji. 9: 181. doi:10.1186/1471-2148-9-181. PMC 2728732. PMID 19650893., see table
- ^ Görmek the remarks of genetic genealogist Robert Tarín for example. We can add 6.1 percent (eight out of 132) in Küba
- ^ Mendizabal, I.; Sandoval, K.; Berniell-Lee, G.; Calafell, F.; Salas, A.; Martinez-Fuentes, A.; Comas, D. (2008). "Küba'daki anne ve babaya ait insan soylarında genetik köken, karışım ve asimetri". BMC Evrimsel Biyoloji. 8: 213. doi:10.1186/1471-2148-8-213. PMC 2492877. PMID 18644108. "The presence of chromosomes of North African origin (E3b1b-M81; Cruciani et al., 2004) can also be explained by a Portuguese-mediated influx, since this haplogroup reaches a frequency of 5.6 percent in Portugal (Beleza et al., 2006), quite similar to the frequency found in Rio de Janeiro (5.4 percent) among European contributors."
- ^ Silva, D. A.; Carvalho, E.; Costa, G.; Tavares, L. G.; Amorim, A. N.; Gusmão, L. (2006). "Y-chromosome genetic variation in Rio De Janeiro population". Amerikan İnsan Biyolojisi Dergisi. 18 (6): 829–837. doi:10.1002 / ajhb.20567. PMID 17039481. S2CID 23778828.
- ^ Coco, C.; Magistrelli, P.; Granone, P.; Roncolini, G.; Picciocchi, A. (1992). "Conservative surgery for early cancer of the distal rectum". Diseases of the Colon & Rectum. 35 (2): 131–6. doi:10.1007/BF02050667. PMID 1735314.
| Sub-topics | |
|---|---|
| Genetik tarih Bölgeye göre | |
| Popülasyon genetiği gruba göre | |
| |