Evrimde yatay gen transferi - Horizontal gene transfer in evolution

Evrim tarihini yeniden inşa etmeye çalışan bilim adamlarına, genlerin bazen uzaktaki dallar arasında transfer olabileceği gerçeğiyle meydan okundu. hayat Ağacı. Genlerin bu hareketi, yatay gen transferi (HGT), biyologların verileri yeniden yapılandırmak için güvendikleri bilgileri karıştırarak soyoluş organizmaların. Tersine, HGT, bilim adamlarının hayat ağacını yeniden inşa etmelerine ve tarihlendirmelerine de yardımcı olabilir. Gerçekte, bir gen transferi filogenetik bir işaret olarak veya verici ve alıcı organizmaların çağdaşlığının kanıtı olarak ve soyu tükenmiş biyolojik çeşitliliğin bir izi olarak kullanılabilir.
HGT çok nadiren gerçekleşir - bireysel organizma düzeyinde, böyle bir olayın meydana gelmesi son derece olası değildir. Bununla birlikte, evrimsel tarihin daha büyük ölçeğinde, bu olaylar belirli bir düzen içinde gerçekleşir. Bir yandan bu, biyologları yaşam tarihi için iyi belirteçler olarak bireysel genlerin kullanımını terk etmeye zorlar. Öte yandan, bu geçmiş hakkında neredeyse hiç kullanılmamış büyük bir bilgi kaynağı sağlar.
Yaşamın üç alanı
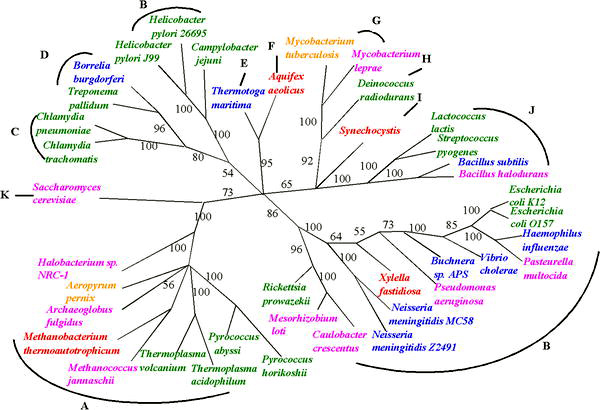
Hayat ağacının üç ana erken dalı yoğun bir şekilde incelenmiştir. mikrobiyologlar çünkü ilk organizmalar mikroorganizmalardı. Mikrobiyologlar (önderlik eden Carl Woese ) terimi tanıttı alan adı bu ağacın üç ana dalı için alan adı anlam olarak biyolojik ile benzer bir filogenetik terimdir krallık. Bu hayat ağacını yeniden inşa etmek için, küçük alt birimini kodlayan gen dizisi ribozomal RNA (SSU rRNA, 16 sn rRNA ) yararlı olduğunu kanıtladı ve ağaç (resimde gösterildiği gibi), bu tek genden gelen bilgilere büyük ölçüde güveniyor.
Bu üç yaşam alanı, erken hücresel yaşamın ana evrimsel soylarını temsil eder ve şu anda şunları içerir: Bakteri, Archaea (yüzeysel olarak bakterilere benzeyen tek hücreli organizmalar) ve Ökarya. Ökarya, yalnızca iyi tanımlanmış bir çekirdeğe sahip organizmaları içerir. mantarlar, protistler ve bitki ve hayvanlar alemindeki tüm organizmalar (şekle bakın).
İçinde filogenetik ilişkiler kurmak için en yaygın olarak kullanılan gen mikroorganizmalar küçük alt birim ribozomal RNA genidir, çünkü dizileri yakın filogenetik mesafeleri olan üyeler arasında korunma eğilimindedir, ancak farklılıkların ölçülebileceği kadar değişken.[1]Evrimsel mesafelerin bir ölçüsü olarak SSU rRNA'nın öncülüğünü, Carl Woese ilk modern "hayat ağacı" nı formüle ederken ve elde ettiği sonuçlar onu, Archaea üçüncü bir alan olarak hayat. Bununla birlikte, son zamanlarda SSU rRNA genlerinin yatay olarak da aktarılabileceği tartışılmıştır.[2] Bu nadir de olsa, bu olasılık, SSU rRNA'lara dayalı olarak filogenetik ağaçların geçerliliğini incelemeye zorluyor.
Mikroorganizmalardaki "yaygın" HGT'nin son keşifleri ve ribozomal RNA'nın küçük alt birimi için eşit genlerin yatay hareketinin tespiti, biyologları ağaçtaki en azından erken dalların doğruluğunu sorgulamaya ve hatta geçerliliğini sorgulamaya zorladı. ağaçlar, evrimin nasıl erken gerçekleştiğine dair yararlı modeller.[3] Aslında, erken evrimin bir topluluktan başlayarak gerçekleştiği kabul edilir. nesiller, HGT standart olduğunda büyük molekülleri değiştirebilir. Bu yanal gen transferi aynı zamanda Darwin eşiği kalıtım veya dikey gen transferi kurulduktan sonra.[4][5]
"Sıra karşılaştırmaları, birçok genler çeşitli arasında Türler dahil olmak üzere filogenetik "alanlar". Dolayısıyla, bir türün filogenetik tarihinin belirlenmesi, tek genler için evrim ağaçlarının belirlenmesiyle kesin olarak yapılamaz. " [6] HGT bu nedenle bir potansiyeldir karışık durum, karıştırıcı faktör çıkarımda filogenetik ağaçlar -den sıra birinin gen. Örneğin, iki uzaktan akraba bakteri bir gen değiştirmişse, filogenetik ağaç bu türlerin dahil edilmesi, diğer genlerin çoğu önemli ölçüde farklılaşmış olsa bile, onların yakından ilişkili olduğunu gösterecektir. Bu nedenle, genlerin varlığı veya yokluğu gibi filogenileri çıkarmak için diğer bilgileri kullanmak veya daha genel olarak, analiz için mümkün olduğunca geniş bir gen yelpazesini dahil etmek önemlidir.
Metafor seçimi: ağaç, ağ, örümcek ağı veya halka
Makalesinde Hayat Ağacını Sökmek, W. Ford Doolittle tartışır Son Evrensel Ortak Ata - Hayat Ağacının kökü - ve bu kavramla ilgili HGT'nin ortaya koyduğu sorunlar.[7] Mikroorganizmayı tarif ediyor Archaeoglobus fulgidus bir anomali olarak filogenetik için kodu temel alan ağaç enzim HMGCoA redüktaz - organizma, bir archaean'dan beklenen tüm hücre lipidleri ve transkripsiyon mekanizmalarıyla kesinlikle bir archaean'dır, ancak HMGCoA genleri bakteri kökenlidir. Makalede Doolittle, artık geniş çapta kabul görürken mitokondri alfa-proteobakteriyel hücrelerden türetilen ökaryotlarda ve kloroplastlar yutulmuştan geldi siyanobakteriler,
".. bunların ilk ökaryotların ortaya çıkmasından sonra meydana gelen tek lateral gen transferleri olduğunu varsaymak artık güvenli değil. Ancak daha sonra, çok hücreli ökaryotlar, yatay gen alışverişi üzerindeki kesin kısıtlamaları biliyoruz, örneğin ayrılmışların gelişi ( ve korumalı) germ hücreleri...
Hiç bir lateral gen transferi olmasaydı, tüm bu bireysel gen ağaçları aynı topolojiye (aynı dallanma sırası) sahip olacak ve her ağacın kökündeki ata genlerinin tümü son evrensel ortak atada mevcut olacaktı. tek antik hücre. Ancak kapsamlı aktarım, durumun böyle olmadığı anlamına gelir: gen ağaçlarının farklı olacağı (birçoğu benzer topolojiye sahip bölgelere sahip olsa da) ve son evrensel ortak ata olarak adlandırılabilecek tek bir hücre asla olmazdı ... "
Doolittle, evrensel ortak atanın belirli bir organizma olamayacağını, ancak birlikte evrimleşmiş ilkel hücrelerin gevşek, çeşitli bir kümelenmesi olması gerektiğini öne sürdü. Her biri nispeten az gene sahip olan bu erken hücreler, birçok yönden farklıydı ve yerlerini değiştirdiler. genler özgürce. Sonunda, bu eklektik hücrelerden bugün bildiğimiz üç yaşam alanı geldi: bakteri, Archaea ve ökaryot. Bu alanlar artık fark edilir şekilde farklıdır çünkü hala meydana gelen gen transferinin çoğu, aralarında değil, bu alanlar içindedir. Biyolog Peter Gogarten bu argümanları pekiştirdi ve bir ağaç metaforunun son genom araştırmalarından elde edilen verilere uymadığını ve bunun yerine biyologların "bir mozaik metaforunu bireysel genomlarda birleştirilmiş farklı geçmişleri tanımlamak için kullanmaları ve [ HGT'nin mikroplar arasındaki zengin alışverişini ve işbirlikçi etkilerini görselleştirmek için bir ağ. "[8]
Filogenomik ile belirsizliğin çözümü
Hayatın başlangıcına kadar filogenilerin yeniden yapılandırılmasındaki belirsizliklere rağmen, HGT'nin ortaya çıkardığı belirsizlikler karşısında hayat ağacının yeniden yapılandırılmasında ilerleme kaydedilmektedir. Tek bir gene dayalı olarak çıkarılan herhangi bir filogenetik ağacın belirsizliği, birkaç ortak gen veya hatta tüm genomlardan elde edilen kanıtlar kullanılarak çözülebilir.[10] Bazen 'çok lokuslu tipleme' olarak adlandırılan bu tür bir yaklaşım, menenjit bakterileri gibi genleri değiştiren organizmalar için filogenik ağaçları çıkarmak için kullanılmıştır.[11]
Jonathan Eisen ve Claire Fraser şunu belirttiler:
"Hayat ağacını oluştururken, tüm genomların analizi, daha önce bir veya birkaç genle yapılan çalışmalar tamamlamaya ve bazı durumlarda iyileştirmeye başladı. Örneğin, tam bakteri genomları üzerine yapılan son araştırmalar, hipertermofilik türlerin derin dallara ayrılmaz; bu doğruysa, yaşamın ilk biçimlerinin termofiller olduğu fikrine şüphe uyandırır.Ökaryotik parazitin genomunun analizi Encephalitozoon cuniculi grubun önerilerini destekler Mikrosporidya derin dallara ayrılan protistler değil, aslında mantar krallığının üyeleridir. Genom analizi, şarbon veya tüberküloza neden olan bakterilerdeki popülasyon genetiği çalışmaları için yeni genetik belirteçler sağlayarak, türler içindeki ilişkileri çözmeye bile yardımcı olabilir. Tüm bu çalışmalarda, filogenetik sinyalin gürültüden ayrılmasına izin veren, eksiksiz bir genom dizisinin sağladığı ek verilerdir. Bu, hayat ağacının artık çözüldüğü anlamına gelmiyor - sadece bir dizi genomu örnekledik ve birçok gruba henüz dokunulmadı "[12]
Bu yaklaşımlar, göreceli HGT sıklığı tahminlerine olanak sağlamaktadır; Gözlemlenen görece düşük değerler, 'ağacın' evrim için hala geçerli bir metafor olduğunu göstermektedir - ancak ağaç yatay olarak aktarılan genlerin 'örümcek ağları' ile süslenmiştir. Bu, Fan Ge, Li-San Wang ve 40'tan fazla tam mikrobiyal genomik dizinin 2005 yılında yapılan bir çalışmasının ana sonucudur. Junhyong Kim. HGT olaylarının sıklığını genom başına çekirdek genlerin yaklaşık% 2'sinde tahmin ediyorlar.[13] Evrimi değerlendirmeye yönelik benzer tüm genom yaklaşımları, ökaryotların iki tam ancak çok çeşitli prokaryot genomunun birleşmesiyle ortaya çıktığı bir öneri gibi, hayat ağacındaki çok erken olayları tanımlamada ilerleme sağlar: biri bir bakteri ve diğeri bir arkay hücresinden. .[3]
Böyle bir organizma füzyonu hipotez karmaşık çekirdekli hücrelerin kökeni için ortaya konmuştur. Lynn Margulis hakkında oldukça farklı akıl yürütme ortakyaşam bir bakteri ve eski bir mikrop konsorsiyumunda ortaya çıkan bir archaen arasında.[14]
Evrimsel çalışmalar için yatay transferin kullanılması
HGT genellikle hayat ağacının yeniden inşası için bir meydan okuma olarak görülse de, alternatif bir görüş, bunun tersine, yeniden inşası için ek değerli bilgiler sağlamasıdır.
İlk olarak, alıcı organizma için HGT, diğerleri gibi bir DNA mutasyonudur ve bu nedenle, ağacın yeniden yapılandırılmasında ve köklenmesinde modellenebilir ve kullanılabilir.[15]
İkincisi, HGT tarafından gen edinimi alan kişinin donör ile aynı zamanda veya daha sonraki bir zamanda yaşaması gerekir.[16] Sonuç olarak, HGT'deki çeşitlendirmenin zamanlaması hakkında bir bilgi var.[17] Canlı dünyada tarihlendirmenin başlıca olağan kaynağı olan fosil kayıtları, mikrobiyal dünyada HGT'nin bol olduğu yerlerde tam olarak bulunmadığından, bu daha da dikkat çekicidir.
Üçüncüsü, nesli tükenmiş biyoçeşitlilik hakkında bilgi sağlar, çünkü transferler muhtemelen soyu tükenmiş türlerden yapılır.[18]
Ayrıca bakınız
Referanslar
- ^ Woese C, vd. (1990). "Doğal bir organizma sistemine doğru: Archaea, Bacteria ve Eucarya alanları için öneri". PNAS ABD. 87 (12): 4576–9. Bibcode:1990PNAS ... 87.4576W. doi:10.1073 / pnas.87.12.4576. PMC 54159. PMID 2112744.
- Woese C, Fox G (1977). "Prokaryotik alanın filogenetik yapısı: birincil krallıklar". PNAS ABD. 74 (11): 5088–90. Bibcode:1977PNAS ... 74.5088W. doi:10.1073 / pnas.74.11.5088. PMC 432104. PMID 270744.
- ^ Yap, WH; Zhang, Z; Wang, Y (1999). "Aktinomisete Thermomonospora chromogena genomunda farklı rRNA operon türleri bulunur ve tüm bir rRNA operonunun yatay transferinin kanıtı". Bakteriyoloji Dergisi. 181 (17): 5201–9. doi:10.1128 / JB.181.17.5201-5209.1999. PMC 94023. PMID 10464188.
- ^ a b Simonson, AB; Servin, JA; Skophammer, RG; Herbold, CW; Rivera, MC; Göl, JA (2005). "Genomik hayat ağacının kodunu çözmek". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 102 Özel Sayı 1: 6608–13. Bibcode:2005PNAS..102.6608S. doi:10.1073 / pnas.0501996102. PMC 1131872. PMID 15851667.
- ^ Woese, C. (1998-06-09). "Evrensel ata". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 95 (12): 6854–6859. Bibcode:1998PNAS ... 95.6854W. doi:10.1073 / pnas.95.12.6854. ISSN 0027-8424. PMC 22660. PMID 9618502.
- ^ Woese, Carl R. (2002-06-25). "Hücrelerin evrimi üzerine". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 99 (13): 8742–8747. Bibcode:2002PNAS ... 99.8742W. doi:10.1073 / pnas.132266999. ISSN 0027-8424. PMC 124369. PMID 12077305.
- ^ Yatay Gen Transferi, Oklahoma Eyaleti
- ^ Doolittle, WF (2000). "Hayat ağacını sökmek". Sci Am. 282 (2): 90–5. Bibcode:2000SciAm.282b..90D. doi:10.1038 / bilimselamerican0200-90. PMID 10710791.
- ^ Gogarten JP 'Yatay Gen Transferi - Biyoloji İçin Yeni Bir Paradigma' doktora tezi
- Zhaxybayeva, O; Gogarten, JP (2004). "Kladogenez, birleşme ve yaşamın üç alanının evrimi" (PDF). Genetikte Eğilimler. 20 (4): 182–7. doi:10.1016 / j.tig.2004.02.004. PMID 15041172.
- ^ Ge, F .; Wang, L .; Kim, J. (2005). "Yatay gen transferinin genom ölçeğinde yapılan tahminlerle ortaya çıkan yaşamın örümcek ağı". PLOS Biyolojisi. 3 (10): e316. doi:10.1371 / journal.pbio.0030316. PMC 1233574. PMID 16122348.

- ^ Henz, S .; Huson, D .; Auch, A .; Nieselt-Struwe, K .; Schuster, S. (2005). "Tüm genom prokaryotik filogeni". Biyoinformatik. 21 (10): 2329–2335. doi:10.1093 / biyoinformatik / bth324. PMID 15166018.
- Fitzpatrick, DA; Logue, ME; Stajich, JE; Butler, G (2006). "Süper ağaç ve birleşik gen analizinden türetilen 42 tam genoma dayalı bir mantar filogeni". BMC Evrimsel Biyoloji. 6: 99. doi:10.1186/1471-2148-6-99. PMC 1679813. PMID 17121679.
- Fitzpatrick, DA; Logue, ME; Stajich, JE; Butler, G (2006). "Süper ağaç ve birleşik gen analizinden türetilen 42 tam genoma dayalı bir mantar filogeni". BMC Evrimsel Biyoloji. 6: 99. doi:10.1186/1471-2148-6-99. PMC 1679813. PMID 17121679.
- ^ Urwin, R; Kız, MC (2003). "Çok lokuslu dizi tipleme: küresel epidemiyoloji için bir araç". Trend Mikrobiyol. 11 (10): 479–87. doi:10.1016 / j.tim.2003.08.006. PMID 14557031.
- Yang, Z (2002). "Birden fazla lokustan gelen verileri kullanarak hominoidlerdeki ataların popülasyon boyutlarının Olasılık ve Bayes tahmini". Genetik. 162 (4): 1811–23. PMC 1462394. PMID 12524351.
- Jennings, W. B .; Edwards, S.V. (2005). "Avustralya çim ispinozlarının (Poephila) özgün tarihi otuz gen ağacından çıkarsanmıştır". Evrim; Uluslararası Organik Evrim Dergisi. 59 (9): 2033–2047. doi:10.1554/05-280.1. PMID 16261740. S2CID 18665436.
- ^ Eisen, J .; Fraser, C. (2003). "Filogenomik: evrim ve genomiğin kesişimi". Bilim. 300 (5626): 1706–1707. Bibcode:2003Sci ... 300.1706E. doi:10.1126 / science.1086292. PMID 12805538. S2CID 42394233.
- ^ Ge, Fan; Wang, Li-San; Kim, Junhyong (2005). "Yatay Gen Transferinin Genom Ölçekli Tahminleriyle Ortaya Çıkan Yaşam Örümcek Ağı". PLOS Biyolojisi. 3 (10): e316. doi:10.1371 / journal.pbio.0030316. PMC 1233574. PMID 16122348.
- ^ Genomları elde etmek: türlerin kökeni hakkında bir teori. Margulis L ve Sagan D (2002) Temel Kitaplar ISBN 0-465-04392-5
- ^ Abby, Sophie S .; Tannier, Eric; Gouy, Manolo; Daubin Vincent (2012-03-27). "Hayat ağacına destek olarak yanal gen transferi". Ulusal Bilimler Akademisi Bildiriler Kitabı. 109 (13): 4962–4967. Bibcode:2012PNAS..109.4962A. doi:10.1073 / pnas.1116871109. PMC 3323970. PMID 22416123.
- ^ Huang, Jinling; Gogarten, Johann Peter (2009). Yatay Gen Transferi. Moleküler Biyolojide Yöntemler. 532. Humana Press. s. 127–139. doi:10.1007/978-1-60327-853-9_7. ISBN 9781603278522. PMID 19271182.
- ^ Davin, Adrián A .; Tannier, Eric; Williams, Tom A .; Boussau, Bastien; Daubin, Vincent; Szöllősi, Gergely J. (2018/04/02). "Gen transferleri hayat ağacına tarih atabilir". Doğa Ekolojisi ve Evrimi. 2 (5): 904–909. doi:10.1038 / s41559-018-0525-3. ISSN 2397-334X. PMC 5912509. PMID 29610471.
- ^ Szöllősi, Gergely J .; Tannier, Eric; Lartillot, Nicolas; Daubin Vincent (2013-05-01). "Ölülerden Yanal Gen Transferi". Sistematik Biyoloji. 62 (3): 386–397. arXiv:1211.4606. doi:10.1093 / sysbio / syt003. ISSN 1063-5157. PMC 3622898. PMID 23355531.
- Bu makale, Citizendium makale "Prokaryot soyoluşu ve evrimi ", altında lisanslı olan Creative Commons Attribution-ShareAlike 3.0 Unported Lisansı ama altında değil GFDL.
daha fazla okuma
- Gogarten, JP; Townsend, JP (2005). "Yatay gen transferi, genom yeniliği ve evrimi". Doğa Yorumları. Mikrobiyoloji. 3 (9): 679–87. doi:10.1038 / nrmicro1204. PMID 16138096. S2CID 8092665.. Derginin tüm sayısında bir makale Doğa İncelemeleri Mikrobiyoloji büyük ölçüde HGT'ye adanmıştır.
- Daubin, V; Moran, NA; Ochman, H (2003). "Filogenetik ve bakteri genomlarının kohezyonu". Bilim. 301 (5634): 829–32. Bibcode:2003Sci ... 301..829D. doi:10.1126 / science.1086568. PMID 12907801. S2CID 11268678.
- Eisen, JA; Fraser, CM (2003). "Bakış açısı filogenomiği: evrim ve genomiğin kesişimi". Bilim. 300 (5626): 1706–7. Bibcode:2003Sci ... 300.1706E. doi:10.1126 / science.1086292. PMID 12805538. S2CID 42394233.
- Ge, F; Wang, LS; Kim, J (2005). "Yatay gen transferinin genom ölçeğinde yapılan tahminlerle ortaya çıkan Yaşam Örümcek Ağı". PLOS Biyolojisi. 3 (10): e316. doi:10.1371 / journal.pbio.0030316. PMC 1233574. PMID 16122348.
- Henz, SR; Huson, DH; Auch, AF; Nieselt-Struwe, K; Schuster, SC (2005). "Tüm genom prokaryotik filogeni". Biyoinformatik. 21 (10): 2329–35. doi:10.1093 / biyoinformatik / bth324. PMID 15166018.
- Lerat, E; Daubin, V; Ochman, H; Moran, NA (2005). "Bakterilerdeki genomik repertuarların evrimsel kökenleri". PLOS Biyolojisi. 3 (5): e130. doi:10.1371 / journal.pbio.0030130. PMC 1073693. PMID 15799709.
- Urwin, R; Kız, MC (2003). "Çok lokuslu dizi tipleme: küresel epidemiyoloji için bir araç". Trend Mikrobiyol. 11 (10): 479–87. doi:10.1016 / j.tim.2003.08.006. PMID 14557031.
- Adoutte, A; Balavoine, G; Lartillot, N; Lespinet, O; Prud'homme, B; De Rosa, R (2000). "Yeni hayvan soyoluşu: güvenilirlik ve çıkarımlar". Proc Natl Acad Sci ABD. 97 (9): 4453–6. Bibcode:2000PNAS ... 97.4453A. doi:10.1073 / pnas.97.9.4453. PMC 34321. PMID 10781043.
- Delsuc, F; Brinkmann, H; Philippe, H (2005). "ve diğerleri.(2005) Filogenomik ve hayat ağacının yeniden inşası ". Nat Rev Genet. 6 (5): 361–75. CiteSeerX 10.1.1.333.1615. doi:10.1038 / nrg1603. PMID 15861208. S2CID 16379422.
- Steenkamp, ET; Wright, J; Baldauf, SL (2006). "Hayvanların ve mantarların protistan kökenleri". Mol Biol Evol. 23 (1): 93–106. doi:10.1093 / molbev / msj011. PMID 16151185.
- Thomas, CM; Nielsen, KM. (2005). "Bakteriler arasında yatay gen transferinin mekanizmaları ve engelleri". Nat Rev Microbiol. 3 (9): 711–21. doi:10.1038 / nrmicro1234. PMID 16138099. S2CID 1231127.