N-Formilmetiyonin-lösil-fenilalanin - N-Formylmethionine-leucyl-phenylalanine
 | |
| İsimler | |
|---|---|
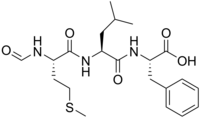
| IUPAC adı (2S)-2-[[(2S)-2-[[(2S) -2-Formamido-4-metilsülfanilbütanoil] amino] -4-metilpentanoil] amino] -3-fenilpropanoik asit | |
| Diğer isimler | |
| Tanımlayıcılar | |
3 boyutlu model (JSmol ) | |
| ChemSpider | |
| MeSH | N-Formilmetiyonin + Leucyl-Fenilalanin |
PubChem Müşteri Kimliği | |
| UNII | |
CompTox Kontrol Paneli (EPA) | |
| |
| |
| Özellikleri | |
| C21H31N3Ö5S | |
| Molar kütle | 437.56 g · mol−1 |
Aksi belirtilmedikçe, veriler kendi içlerindeki malzemeler için verilmiştir. standart durum (25 ° C'de [77 ° F], 100 kPa). | |
| Bilgi kutusu referansları | |
N-Formilmetiyonil-leusil-fenilalanin (fMLF, fMLP veya N-formil-met-leu-phe) bir N-formile tripeptid ve bazen basitçe şöyle anılır kemotaktik peptid güçlü polimorfonükleer lökosit (PMN) kemotaktik faktör ve aynı zamanda bir makrofaj aktivatör.[2]
fMLF, prototip temsilcisidir. Nbiçimlendirilmiş oligopeptid kemotaktik faktörler ailesi. Bu oligopeptidlerin eylemlerini taklit ettiği veya taklit ettiği bilinmektedir. N(a) doku bakterileri tarafından salgılanan, (b) dolaşımdaki kan lökositlerini belirli bölgelere bağlanarak çeken ve aktive eden formil oligopeptitler. G proteinine bağlı reseptörler ve (c) böylelikle enflamatuar yanıtı bakteriyel istila bölgelerine yönlendirir. fMLF, doğuştan gelen bağışıklık ev sahibi savunma mekanizması patojenler.
fMLF, bir kemotaktik faktör için bir lökosit reseptörünün ilk keşfine yol açtı, enflamatuar yanıtlar üzerinde tamamlayıcı ve / veya zıt etkileri olan üç farklı tipte fMLF reseptörü ve diğer birçok aktiviteyi tanımladı ve uyarı-yanıt birleştirme mekanizmalarının tanımlanmasına yardımcı oldu. çeşitli kemotaktik faktörler ve bunların G proteinine bağlı reseptörleri hücresel işlevi indükler.
Keşif
1887'de, Élie Metchnikoff bunu gözlemledim lökositler Çeşitli hayvanların kanından izole edilen bazı bakterilere karşı çekilmiştir.[3] Bu çekiciliğin kısa süre sonra bakteriler tarafından salınan çözünür elementlerden kaynaklandığı öne sürüldü [4] (bakınız Harris[5] 1953'e kadar bu alanın gözden geçirilmesi için). Peter Ward, Elmer Becker, Henry Showell ve meslektaşları, bu unsurların çeşitli büyüme yöntemleriyle yapıldığını gösterdi. gram pozitif bakteriler ve gram negatif bakteriler ve düşüktü moleküler ağırlık yani 3600'ün altında Dalton (birim) s.[6][7][8] Schiffmann ve kolejler tarafından yapılan diğer araştırmalar, büyüyen kültürlerin Escherichia coli 150 ila 1500 dalton arasında serbest bırakılan oligopeptidler Karboksilik asit grup ama bedava değil Amin grubu.[9]
Bakterilerin kopyaladığı bu ipuçları ve bilgiler göz önüne alındığında (bkz. Transkripsiyon (genetik) ) ile başlayan proteinler N-formilmetiyonin ökaryotik hücreler çoğunlukla formile edilmemiş metiyonin ile protein sentezini başlatırken, Schiffmann, Corcoran ve Wahl teorize edildi ve sonra gösterdi N-formil-metiyonin ve bir dizi N-formil-metiyonil dipeptidler ve tripeptidler, kemotaksisini uyarmıştır. nötrofiller kobay peritoneal eksüdalarından izole edilen makrofajların yanı sıra tavşan peritoneal eksüdalarından izole edilmiştir.[10] Daha ileri çalışmalarda çeşitli N-formillenmiş oligopeptitler, fMLF, tavşan nötrofil kemotaksisini uyarmada en güçlü olduğunu kanıtladı.[7] fMLF ve diğer, daha az etkili bir örnekleme, N-formil oligopeptidlerin daha sonra geniş bir yelpazede tavşan nötrofil fonksiyonlarını uyardığı bulundu: bu hücrelerin süspansiyonda geçici olarak otomatik toplanması[11] ve tavşan damarlarına enjekte edildiğinde dolaşımdaki nötrofillerde eşit ölçüde geçici düşüş (bu tepkiler nötrofillerin birbirine ve / veya vasküler endotelyuma yapışmasındaki artıştan kaynaklanır);[12] sürüm (bkz. degranülasyon ) hücre içi granüle bağlı enzimler ve diğer antimikrobiyal sitotoksik moleküller; ve sitotoksik üretim ve salınım Reaktif oksijen türleri gibi Süperoksit ve hidrojen peroksit.[7][13] Tüm bu yanıtlar, doğuştan gelen bağışıklık sistemleri bakteri istilalarına karşı ilk savunma hattı.
Takip çalışmaları, mitokondri ve kloroplast genlerinin organellerinin Ökaryot İnsanlarınki de dahil olmak üzere, nükleer genlerin aksine, N-formil-metiyonin proteinleri, salım NfMLF kemotaksisininkini tam olarak taklit eden kemotaktik aktiviteye sahip peptitler içeren formil-metiyonil[14] Bu organelden türetilmiş formillenmiş peptidler, fMLF reseptörleri aracılığıyla dolaşan kan lökositlerini görevlendirmek ve böylece bakterilerin neden olmadığı hücre hasarı ve doku yıkımı bölgelerinde iltihap tepkilerini başlatan gerçek fMLF analoglarıdır.[15] fMLF ve diğer N-formillenmiş oligopeptidlerin insan nötrofillerinde benzer şekilde aktif olduğu bulunmuştur.[16][17] Bu nötrofil tepkilerini uyarmada geniş bir dizi formillenmiş peptidin yüksek derecede yapısal özgüllüğü, N- uyarıcı güçlerine paralel afinitelere sahip nötrofillere formüle edilmiş oligopeptitler,[18] yeteneği t-karbobenzoksi-fenilalanil-metiyonin nötrofillere bağlanır ancak uyarmaz ve böylece N-formillenmiş oligopeptitlerin nötrofil bağlanmasını ve uyarıcı aktivitesini bloke eder,[19][20] ve formillenmiş oligopeptidlerin nötrofil işlevsel tepkilerini kendilerine duyarsızlaştırma (yani tepkisiz hale getirme) yeteneği, ancak bir dizi başka kemotaktik uyarana karşı duyarsızlaştırma yeteneği yoktur veya daha azdır.[21][22] Formillenmiş peptidlerin, diğer kemotaktik faktör reseptörlerinden farklı olan ortak, özel bir reseptör sistemi aracılığıyla hücreler üzerinde hareket ettiğine dair güçlü düşündürücü kanıtlar sağladı.
Reseptörler
Yukarıda belirtilen çalışmalar, nihai klonlama insanın Formil peptid reseptörü 1, bir G proteinine bağlı reseptör insan ve tavşan nötrofilleri üzerindeki uyarıcı etkilerine aracılık etmek için fMLF ve diğer formüle edilmiş oligopeptitleri bağlayan. Daha sonra Formil peptid reseptörü 2 ve Formil peptid reseptörü 3 Formil peptid reseptörü 1'in amino asit sekansındaki benzerliklere göre de klonlanmıştır. Formil peptid reseptörleri 2 ve 3, formil peptid reseptörü 1 ile karşılaştırıldığında fMLF dahil olmak üzere formillenmiş oligopeptidlere bağlanma ve yanıt verme konusunda çok farklı yeteneklere sahiptir ve birbirleriyle karşılaştırılmıştır. ve formil peptid reseptörü 1'inkinden çok farklı işlevlere sahiptir.[23]
Referanslar
- ^ n-formilmetiyonin lösil-fenilalanin, Cancerweb
- ^ Panaro MA, Mitolo V (Ağu 1999). "FMLF'ye meydan okuyan hücresel yanıtlar: bir mini inceleme". İmmünofarmakoloji ve İmmünotoksikoloji. 21 (3): 397–419. doi:10.3109/08923979909007117. PMID 10466071.
- ^ Metchnikoff E (1887). "Sur la lutte des cellules de l'organisme contre l'invasion des microsbes". Ann. Inst. Pastör. 1: 321.
- ^ Grawitz P (1887). "Bilinmeyen". Virchows Adz. IIO. ben. Alıntı genel başlığı kullanır (Yardım)
- ^ Harris H (Temmuz 1954). "Enflamasyonda kemotaksinin rolü". Fizyolojik İncelemeler. 34 (3): 529–62. doi:10.1152 / physrev.1954.34.3.529. PMID 13185754.
- ^ Ward PA, Lepow IH, Newman LJ (Nisan 1968). "Polimorfonükleer lökositler için kemotaktik bakteriyel faktörler". Amerikan Patoloji Dergisi. 52 (4): 725–36. PMC 2013377. PMID 4384494.
- ^ a b c Showell HJ, Freer RJ, Zigmond SH, Schiffmann E, Aswanikumar S, Corcoran B, Becker EL (Mayıs 1976). "Sentetik peptitlerin, kemotaktik faktörler ve nötrofiller için lizozomal sekresyon indükleyicileri olarak yapı-aktivite ilişkileri". Deneysel Tıp Dergisi. 143 (5): 1154–69. doi:10.1084 / jem.143.5.1154. PMC 2190180. PMID 1262785.
- ^ Becker EL, Showell HJ (Haziran 1974). "Kemotaktik faktörlerin lizozomal enzim salımını indükleme yeteneği. II. Salım mekanizması". Journal of Immunology. 112 (6): 2055–62. PMID 4825785.
- ^ Schiffmann E, Showell HV, Corcoran BA, Ward PA, Smith E, Becker EL (Haziran 1975). "Escherichia coli'den nötrofil kemotaktik faktörlerin izolasyonu ve kısmi karakterizasyonu". Journal of Immunology. 114 (6): 1831–7. PMID 165239.
- ^ Schiffmann E, Corcoran BA, Wahl SM (Mart 1975). "Lökositler için kemoatraktan olarak N-formilmetiyonil peptitler". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 72 (3): 1059–62. doi:10.1073 / pnas.72.3.1059. PMC 432465. PMID 1093163.
- ^ O'Flaherty JT, Kreutzer DL, Ward PA (Temmuz 1977). "Kemotaktik ajanların neden olduğu nötrofil toplanması ve şişmesi". Journal of Immunology. 119 (1): 232–9. PMID 874320.
- ^ O'Flaherty JT, Showell HJ, Ward PA (Mayıs 1977). "Kemotaktik faktörlerin sistemik infüzyonuyla indüklenen nötropeni". Journal of Immunology. 118 (5): 1586–9. PMID 858915.
- ^ Becker EL, Sigman M, Oliver JM (Nisan 1979). "Tavşan polimorfonükleer lökositlerinde sentetik kemotaktik peptitler ve A23187 tarafından indüklenen süperoksit üretimi". Amerikan Patoloji Dergisi. 95 (1): 81–97. PMC 2042294. PMID 219701.
- ^ Sazan H (Ocak 1982). "Nötrofiller için kemoatraktan olarak mitokondriyal N-formilmetiyonil proteinleri". Deneysel Tıp Dergisi. 155 (1): 264–75. doi:10.1084 / jem.155.1.264. PMC 2186576. PMID 6274994.
- ^ Dorward DA, Lucas CD, Chapman GB, Haslett C, Dhaliwal K, Rossi AG (Mayıs 2015). "Akut iltihaplanma sırasında nötrofil işlevini yönetmede formillenmiş peptidlerin ve formil peptid reseptör 1'in rolü". Amerikan Patoloji Dergisi. 185 (5): 1172–84. doi:10.1016 / j.ajpath.2015.01.020. PMC 4419282. PMID 25791526.
- ^ O'Flaherty JT, Kreutzer DL, Ward PA (Mart 1978). "Kemotaktik faktör, insan lökositlerinin topaklanması, şişmesi ve yabancı yüzey yapışkanlığını etkiler". Amerikan Patoloji Dergisi. 90 (3): 537–50. PMC 2018255. PMID 564610.
- ^ Lehmeyer JE, Snyderman R, Johnston RB (Temmuz 1979). "Nötrofil oksidatif metabolizmasının kemotaktik peptidlerle uyarılması: kalsiyum iyon konsantrasyonu ve sitokalasin B'nin etkisi ve forbol miristat asetat ile uyarımla karşılaştırma". Kan. 54 (1): 35–45. doi:10.1182 / blood.V54.1.35.35. PMID 444673.
- ^ Aswanikumar S, Corcoran B, Schiffmann E, Day AR, Freer RJ, Showell HJ, Becker EL (Ocak 1977). "Kemotaktik peptitler için tavşan nötrofillerinde bir reseptörün gösterilmesi". Biyokimyasal ve Biyofiziksel Araştırma İletişimi. 74 (2): 810–7. doi:10.1016 / 0006-291x (77) 90375-8. PMID 836328.
- ^ Schiffmann, E., B. A. Corcoran ve A. Aswanikumar. (1978). "Nötrofillerin sentetik N-formilmetiyonin kemotaktik peptitlere tepkisindeki moleküler olaylar." J. A. Gallin; P. G. Quie (editörler). Lökosit kemotaksisi: Metodoloji, fizyoloji, klinik çıkarımlar. New York .: Raven Press.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ O'Flaherty JT, Showell HJ, Kreutzer DL, Ward PA, Becker EL (Nisan 1978). "Bir rekabetçi antagonist tarafından kemotaktik faktörlere karşı in vivo ve in vitro nötrofil yanıtlarının inhibisyonu". Journal of Immunology. 120 (4): 1326–32. PMID 641351.
- ^ O'Flaherty JT, Kreutzer DL, Showell HS, Becker EL, Ward PA (Aralık 1978). "Kemotaktik faktörlere nötrofil agregasyon yanıtının duyarsızlaşması". Amerikan Patoloji Dergisi. 93 (3): 693–706. PMC 2018345. PMID 717543.
- ^ O'Flaherty JT, Kreutzer DL, Showell HJ, Vitkauskas G, Becker EL, Ward PA (Mart 1979). "Kemotaktik faktörlere karşı seçici nötrofil duyarsızlaştırma". Hücre Biyolojisi Dergisi. 80 (3): 564–72. doi:10.1083 / jcb.80.3.564. PMC 2110355. PMID 457760.
- ^ Li Y, Ye D (Temmuz 2013). "İnsan hastalıklarında formil peptid reseptörleri için moleküler biyoloji". Moleküler Tıp Dergisi. 91 (7): 781–9. doi:10.1007 / s00109-013-1005-5. PMID 23404331. S2CID 150459.
Dış bağlantılar
- N-Formilmetiyonin Leucyl-Fenilalanin ABD Ulusal Tıp Kütüphanesinde Tıbbi Konu Başlıkları (MeSH)